

Tissue-specific dynamic codon redefinition in Drosophila
- PMID: 33500350
- PMCID: PMC7865143
- DOI: 10.1073/pnas.2012793118
Tissue-specific dynamic codon redefinition in Drosophila
Abstract
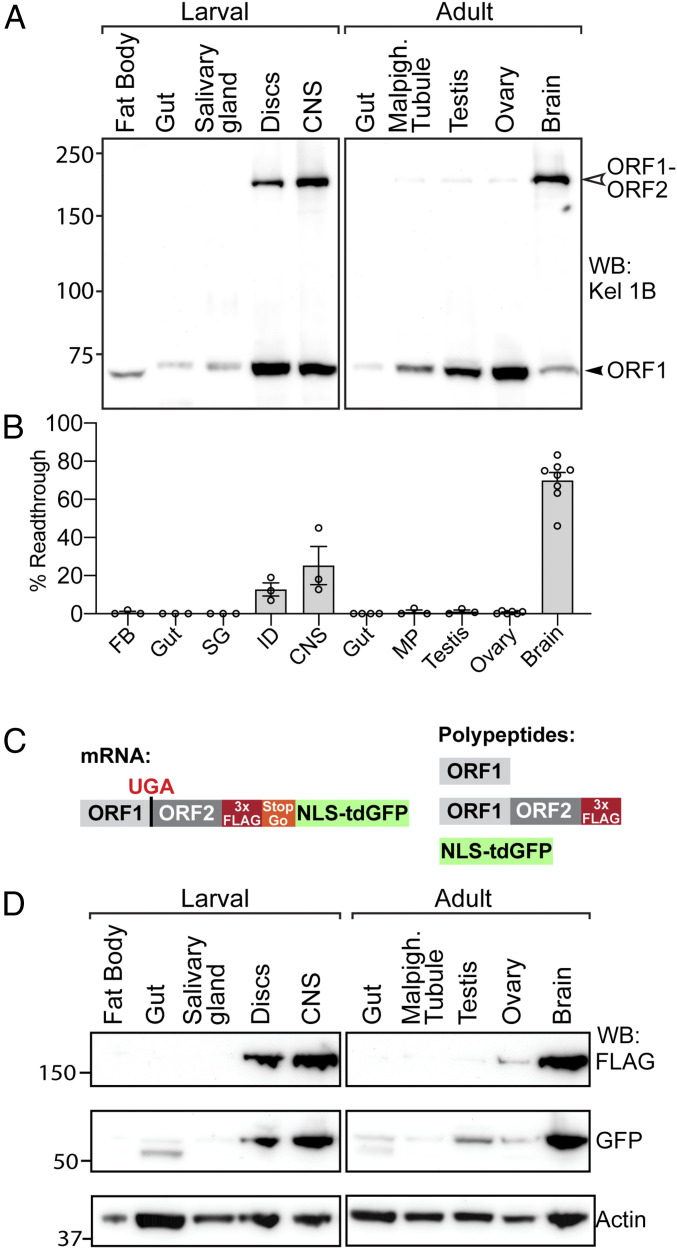
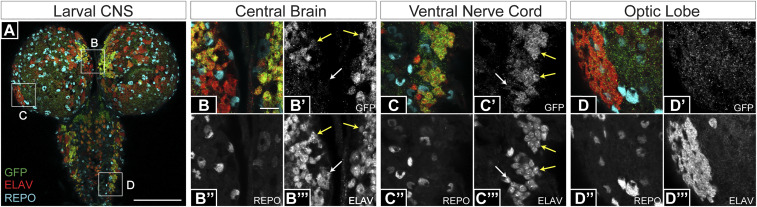
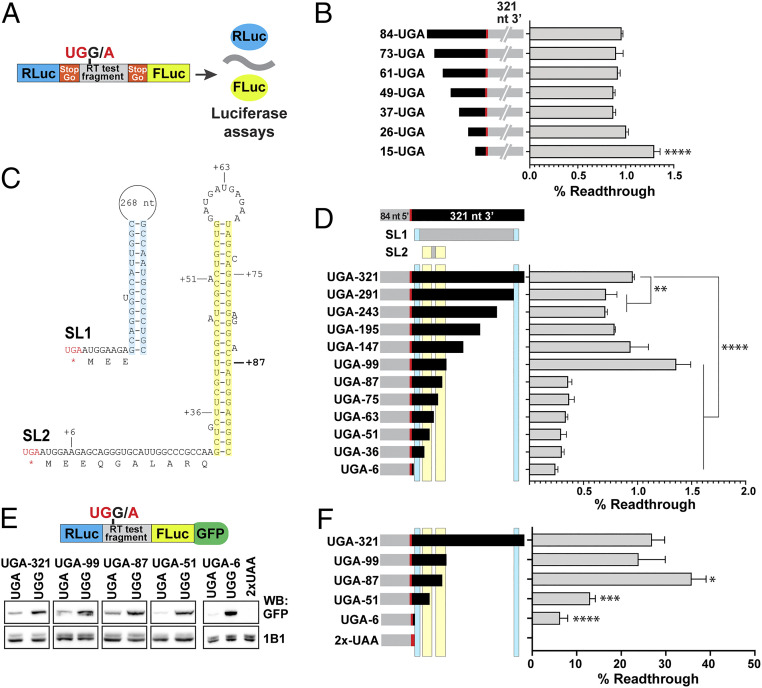
Translational stop codon readthrough occurs in organisms ranging from viruses to mammals and is especially prevalent in decoding Drosophila and viral mRNAs. Recoding of UGA, UAG, or UAA to specify an amino acid allows a proportion of the protein encoded by a single gene to be C-terminally extended. The extended product from Drosophila kelch mRNA is 160 kDa, whereas unextended Kelch protein, a subunit of a Cullin3-RING ubiquitin ligase, is 76 kDa. Previously we reported tissue-specific regulation of readthrough of the first kelch stop codon. Here, we characterize major efficiency differences in a variety of cell types. Immunoblotting revealed low levels of readthrough in malpighian tubules, ovary, and testis but abundant readthrough product in lysates of larval and adult central nervous system (CNS) tissue. Reporters of readthrough demonstrated greater than 30% readthrough in adult brains, and imaging in larval and adult brains showed that readthrough occurred in neurons but not glia. The extent of readthrough stimulatory sequences flanking the readthrough stop codon was assessed in transgenic Drosophila and in human tissue culture cells where inefficient readthrough occurs. A 99-nucleotide sequence with potential to form an mRNA stem-loop 3' of the readthrough stop codon stimulated readthrough efficiency. However, even with just six nucleotides of kelch mRNA sequence 3' of the stop codon, readthrough efficiency only dropped to 6% in adult neurons. Finally, we show that high-efficiency readthrough in the Drosophila CNS is common; for many neuronal proteins, C-terminal extended forms of individual proteins are likely relatively abundant.
Keywords: Drosophila; Kelch; central nervous system (CNS); recoding; stop codon readthrough.
Conflict of interest statement
The authors declare no competing interest.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
