

Influenza Causes MLKL-Driven Cardiac Proteome Remodeling During Convalescence
- PMID: 33501852
- PMCID: PMC8313652
- DOI: 10.1161/CIRCRESAHA.120.318511
Influenza Causes MLKL-Driven Cardiac Proteome Remodeling During Convalescence
Abstract
Rationale: Patients with and without cardiovascular diseases have been shown to be at risk of influenza-mediated cardiac complications. Recent clinical reports support the notion of a direct link between laboratory-confirmed influenza virus infections and adverse cardiac events.
Objective: Define the molecular mechanisms underlying influenza virus-induced cardiac pathogenesis after resolution of pulmonary infection and the role of necroptosis in this process.
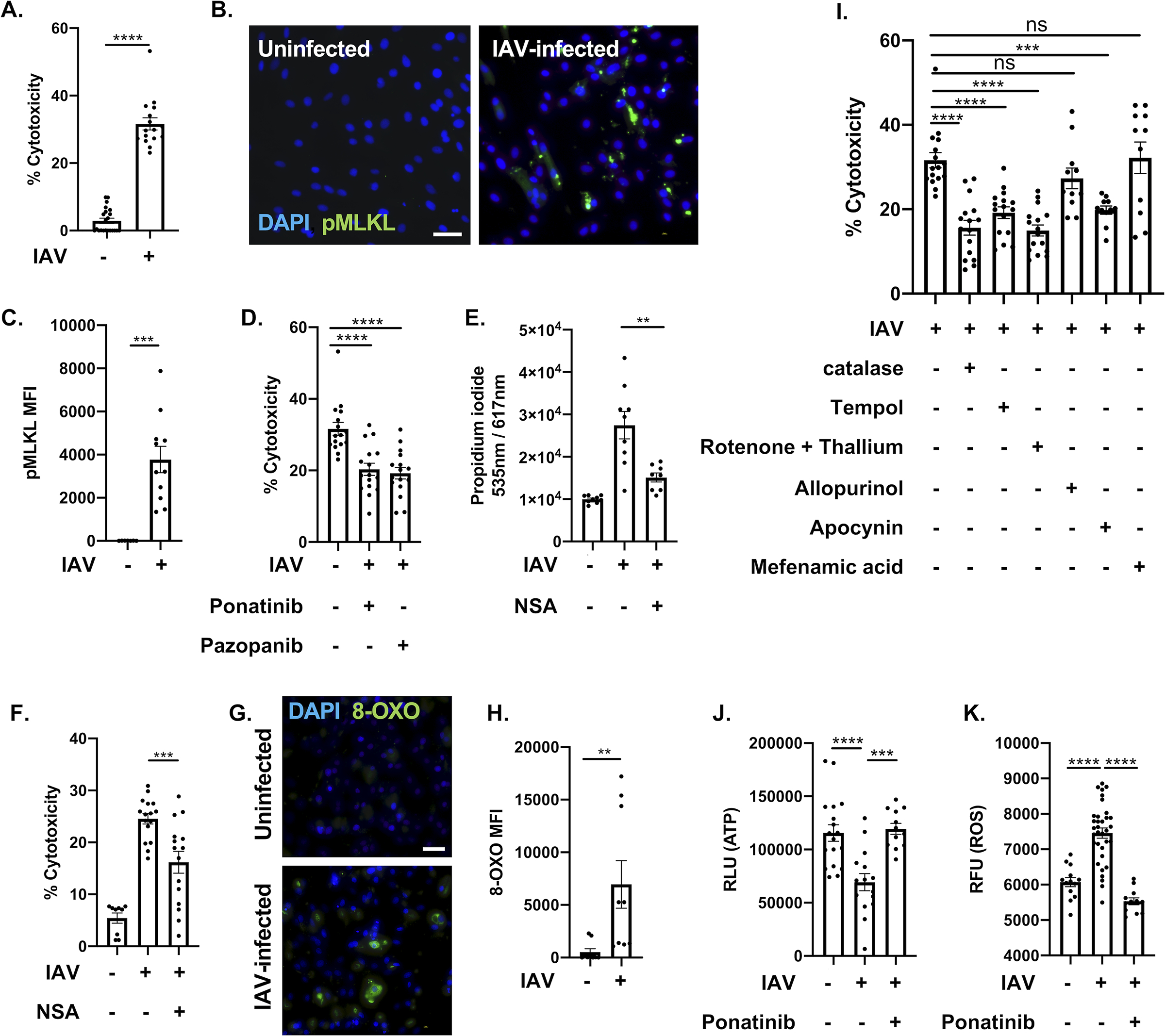
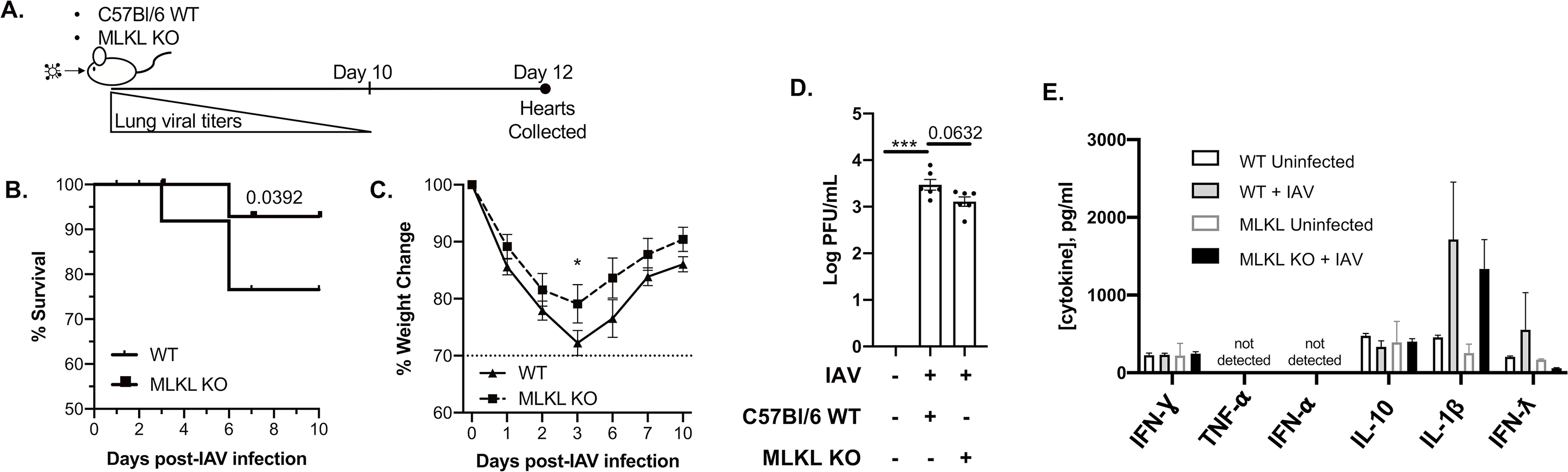
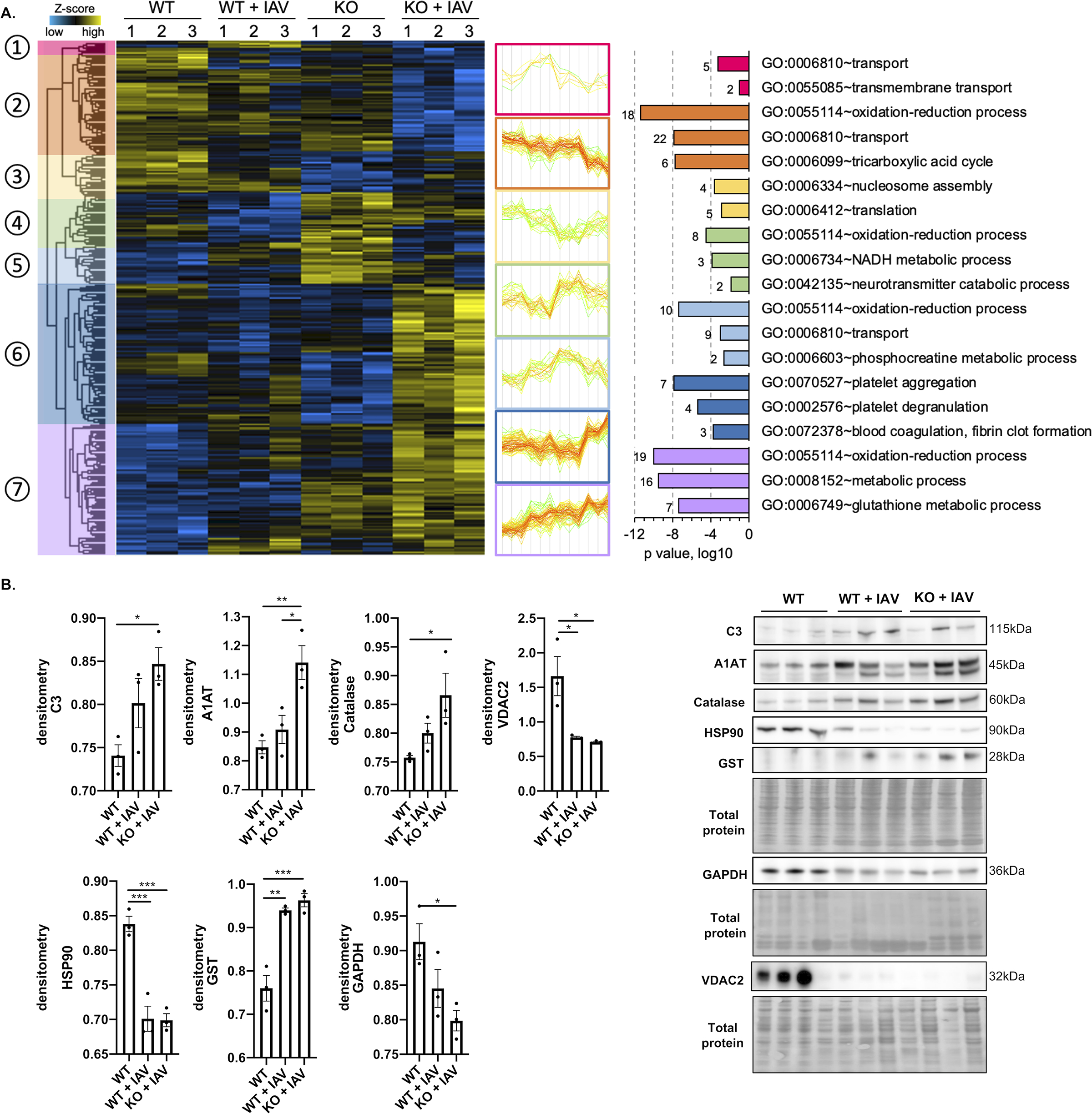
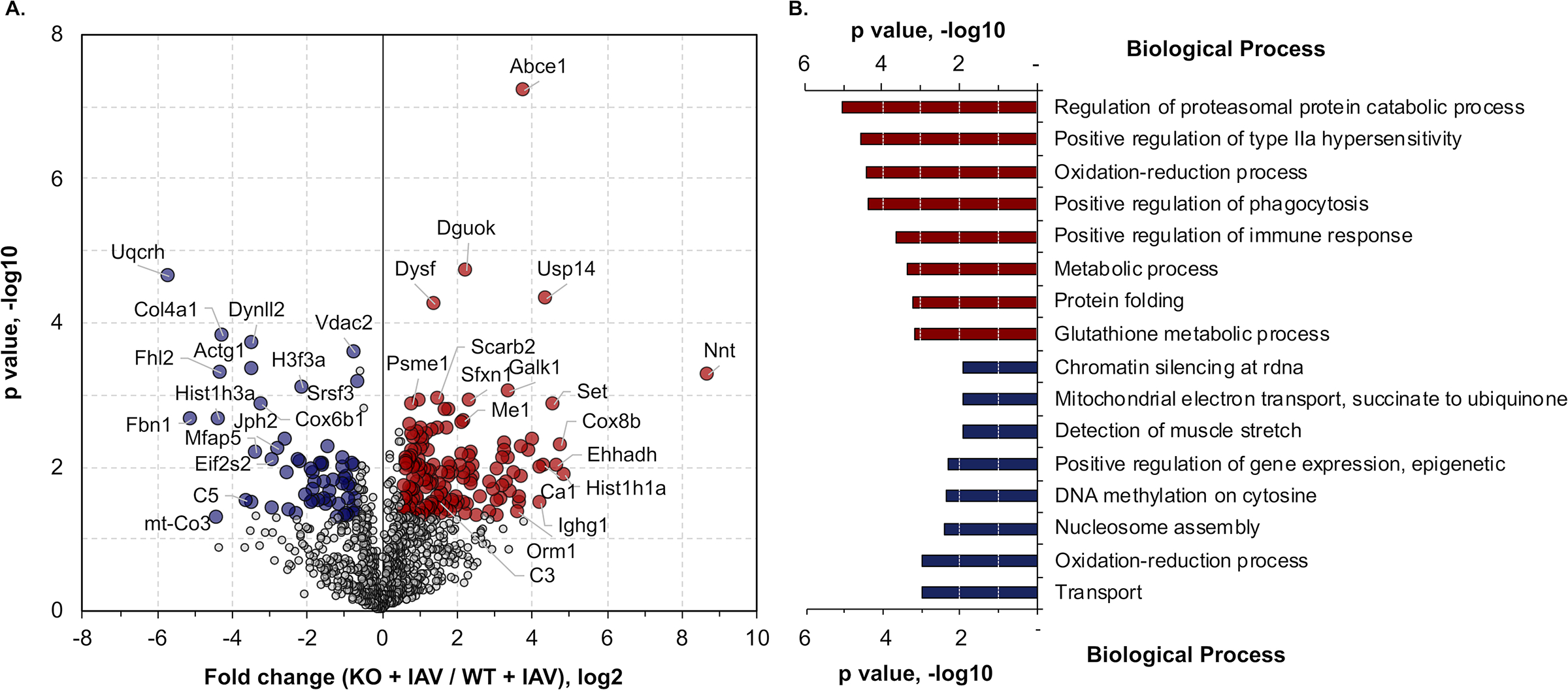
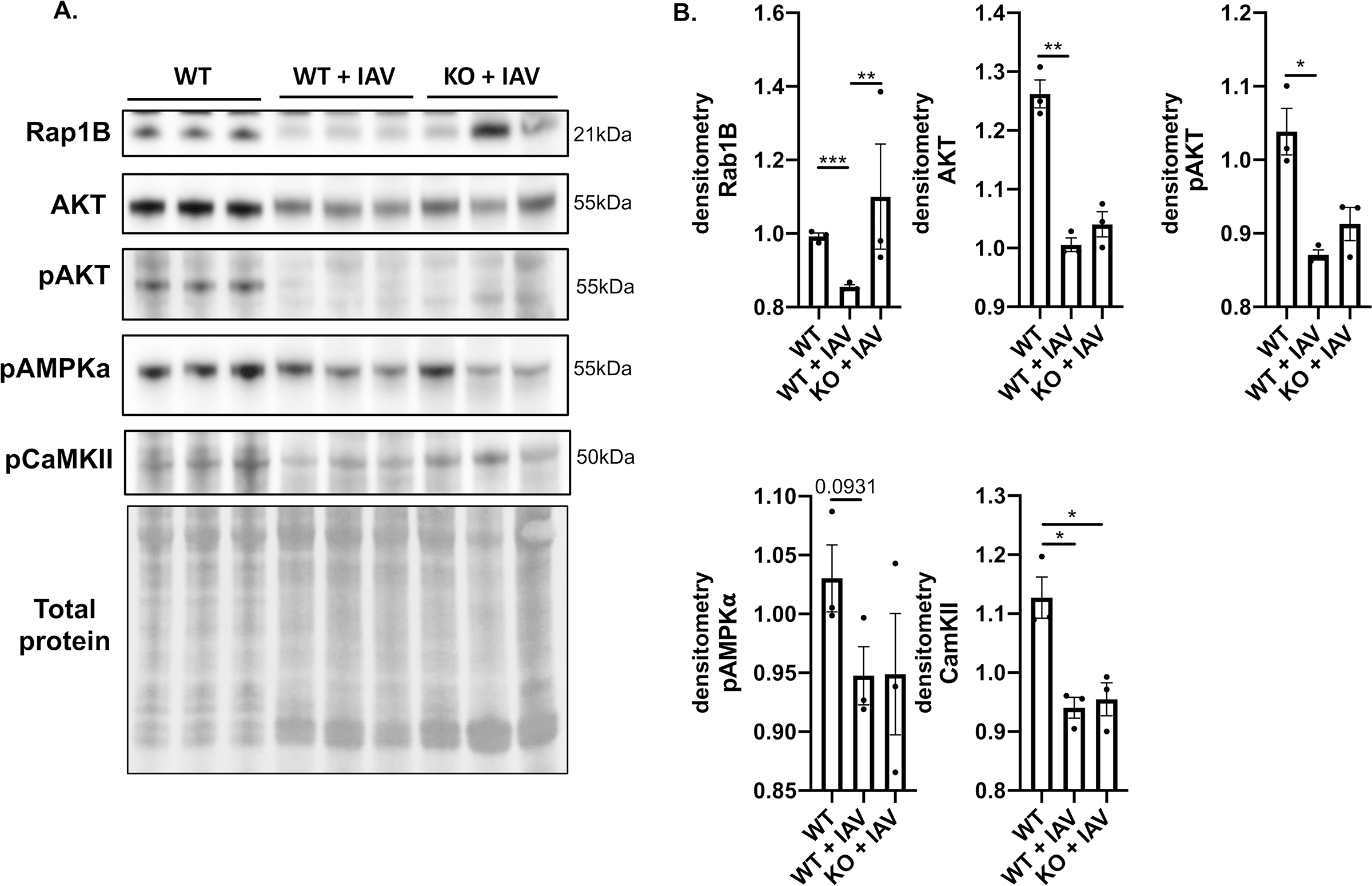
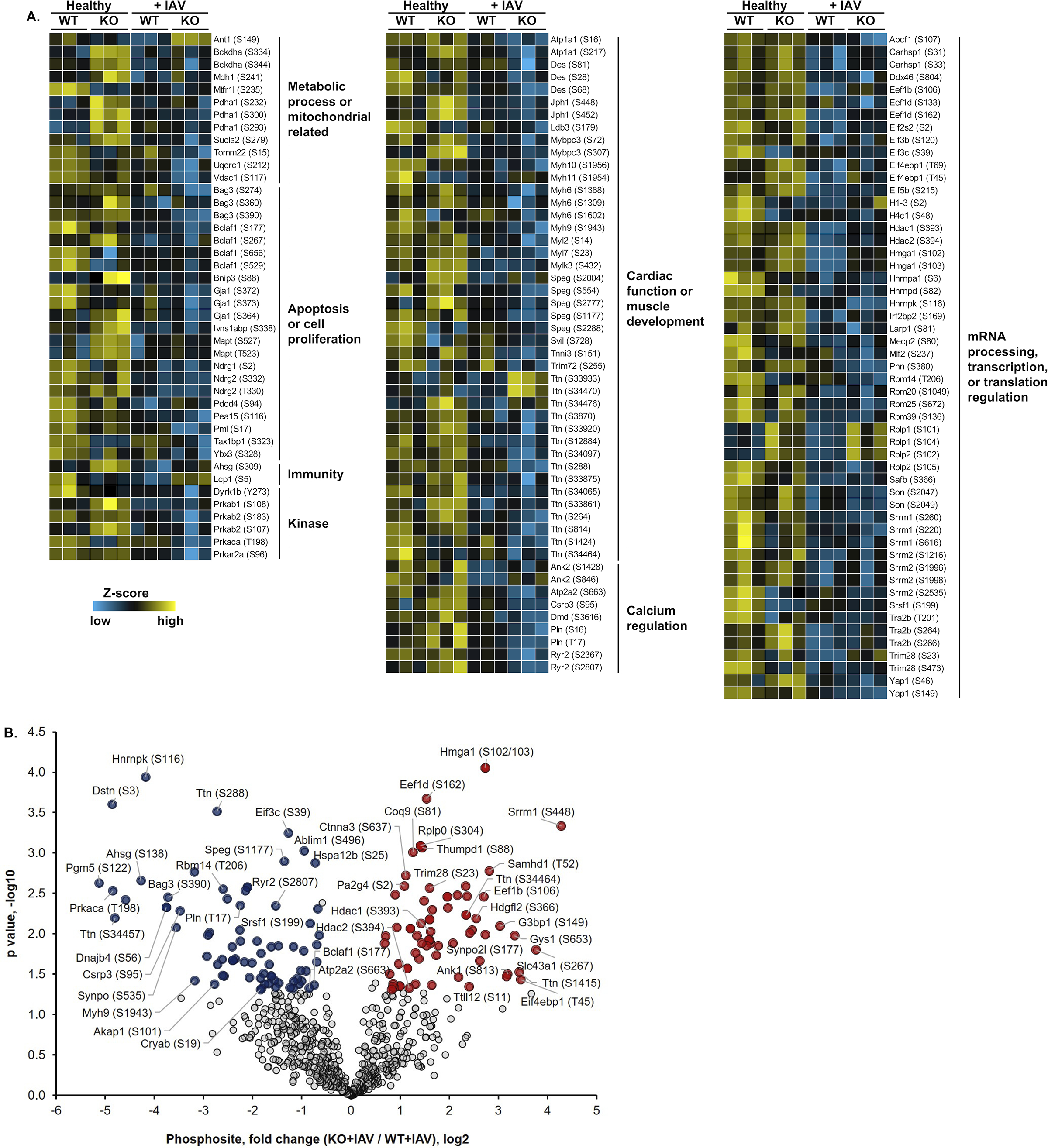
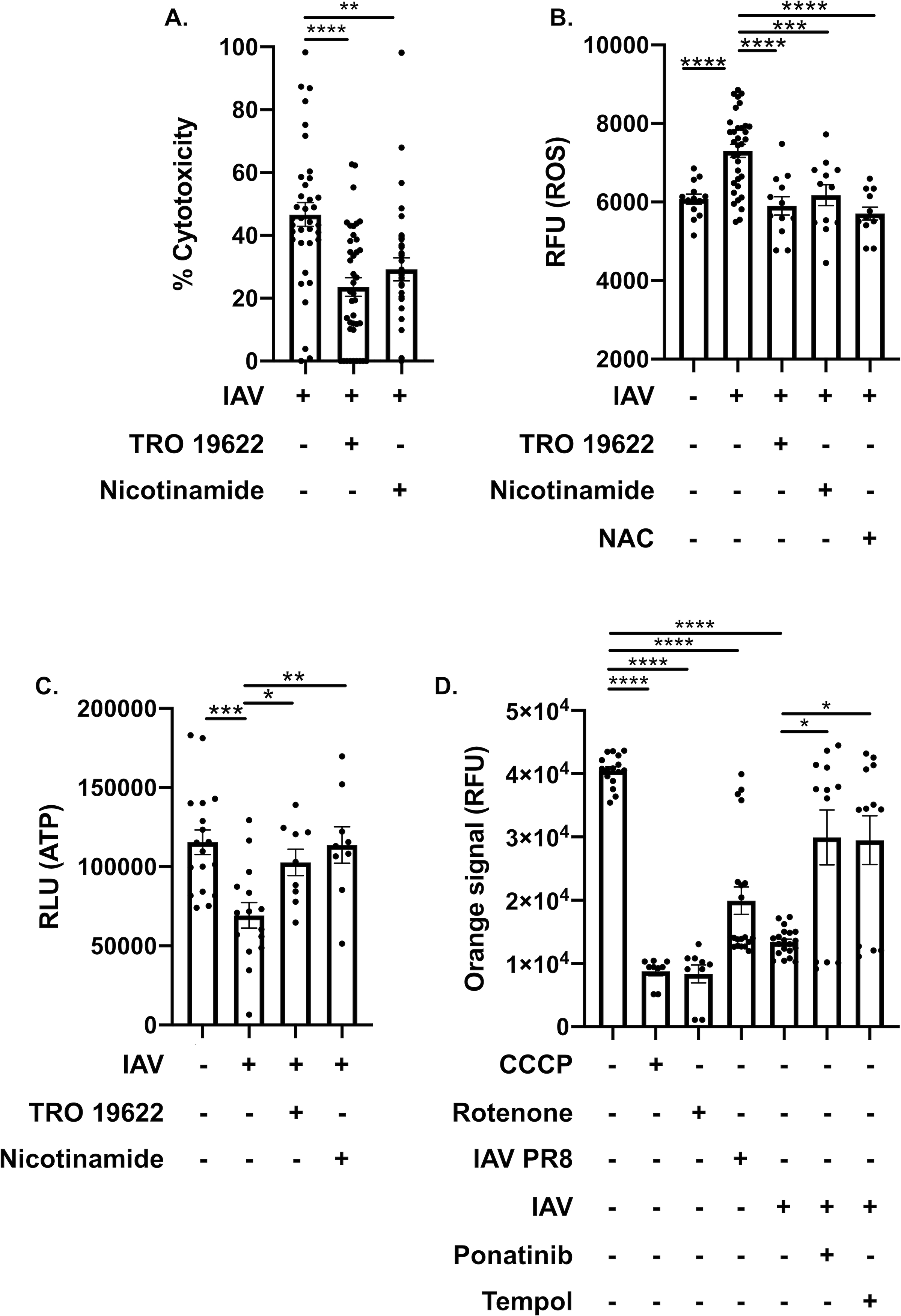
Methods and results: Hearts from wild-type and necroptosis-deficient (MLKL [mixed lineage kinase domain-like protein]-KO) mice were dissected 12 days after initial influenza A virus (IAV) infection when viral titers were undetectable in the lungs. Immunofluorescence microscopy and plaque assays showed presence of viable IAV particles in the myocardium without generation of interferon responses. Global proteome and phosphoproteome analyses using high-resolution accurate mass-based LC-MS/MS and label-free quantitation showed that the global proteome as well as the phosphoproteome profiles were significantly altered in IAV-infected mouse hearts in a strain-independent manner. Necroptosis-deficient mice had increased survival and reduced weight loss post-IAV infection, as well as increased antioxidant and mitochondrial function, indicating partial protection to IAV infection. These findings were confirmed in vitro by pretreatment of human and rat myocytes with antioxidants or necroptosis inhibitors, which blunted oxidative stress and mitochondrial damage after IAV infection.
Conclusions: This study provides the first evidence that the cardiac proteome and phosphoproteome are significantly altered post-pulmonary influenza infection. Moreover, viral particles can persist in the heart after lung clearance, altering mitochondrial function and promoting cell death without active replication and interferon responses. Finally, our findings show inhibition of necroptosis or prevention of mitochondrial damage as possible therapeutic interventions to reduce cardiac damage during influenza infections. Graphic Abstract: A graphic abstract is available for this article.
Keywords: cell death; heart; human influenza; mitochondria; necroptosis; oxidative stress; proteomics.
Figures
References
-
- Heron M Deaths: Leading causes for 2017. National vital statistics reports. 2019;68 - PubMed
-
- Paules C, Subbarao K. Influenza. Lancet. 2017;390:697–708 - PubMed
-
- Noda T [orthomyxoviruses]. Uirusu. 2012;62:219–228 - PubMed
-
- Thompson WW, Shay DK, Weintraub E, Brammer L, Cox N, Anderson LJ, Fukuda K. Mortality associated with influenza and respiratory syncytial virus in the united states. JAMA. 2003;289:179–186 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
