

Functional properties of insect olfactory receptors: ionotropic receptors and odorant receptors
- PMID: 33502604
- PMCID: PMC7873100
- DOI: 10.1007/s00441-020-03363-x
Functional properties of insect olfactory receptors: ionotropic receptors and odorant receptors
Abstract
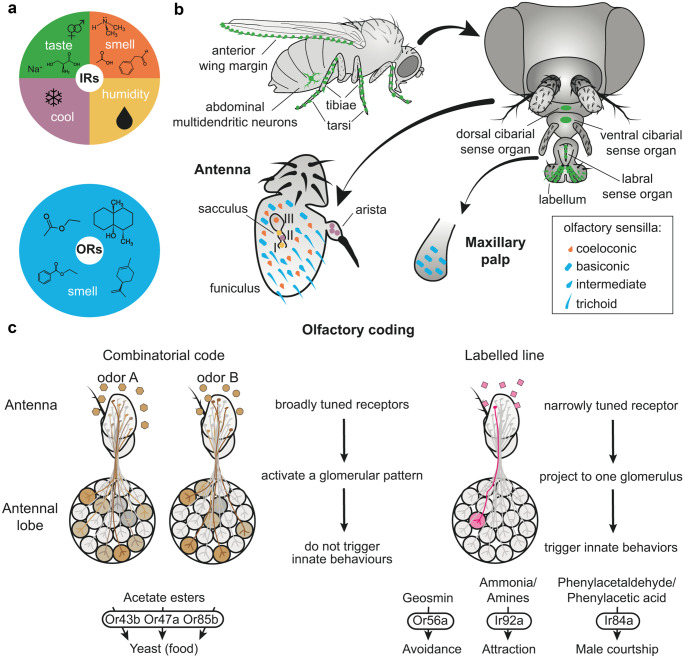
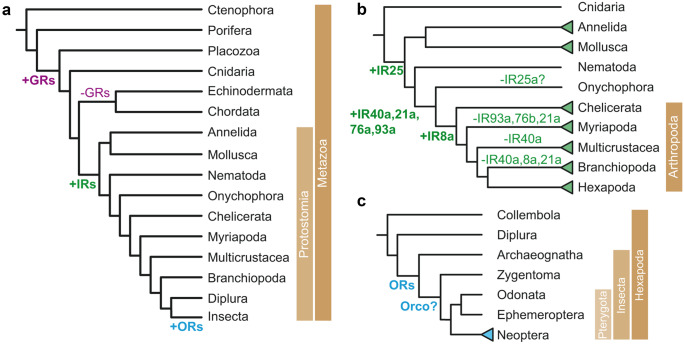
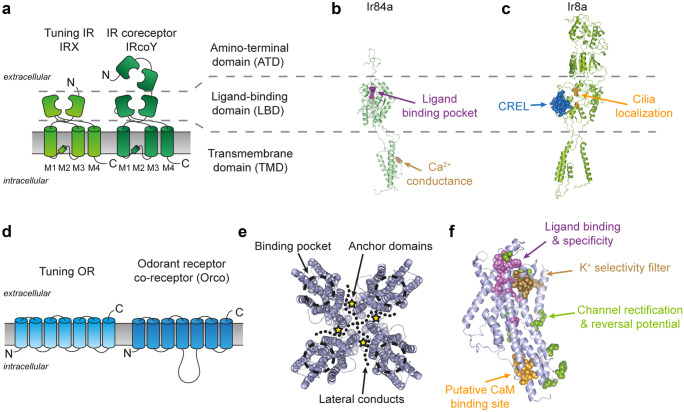
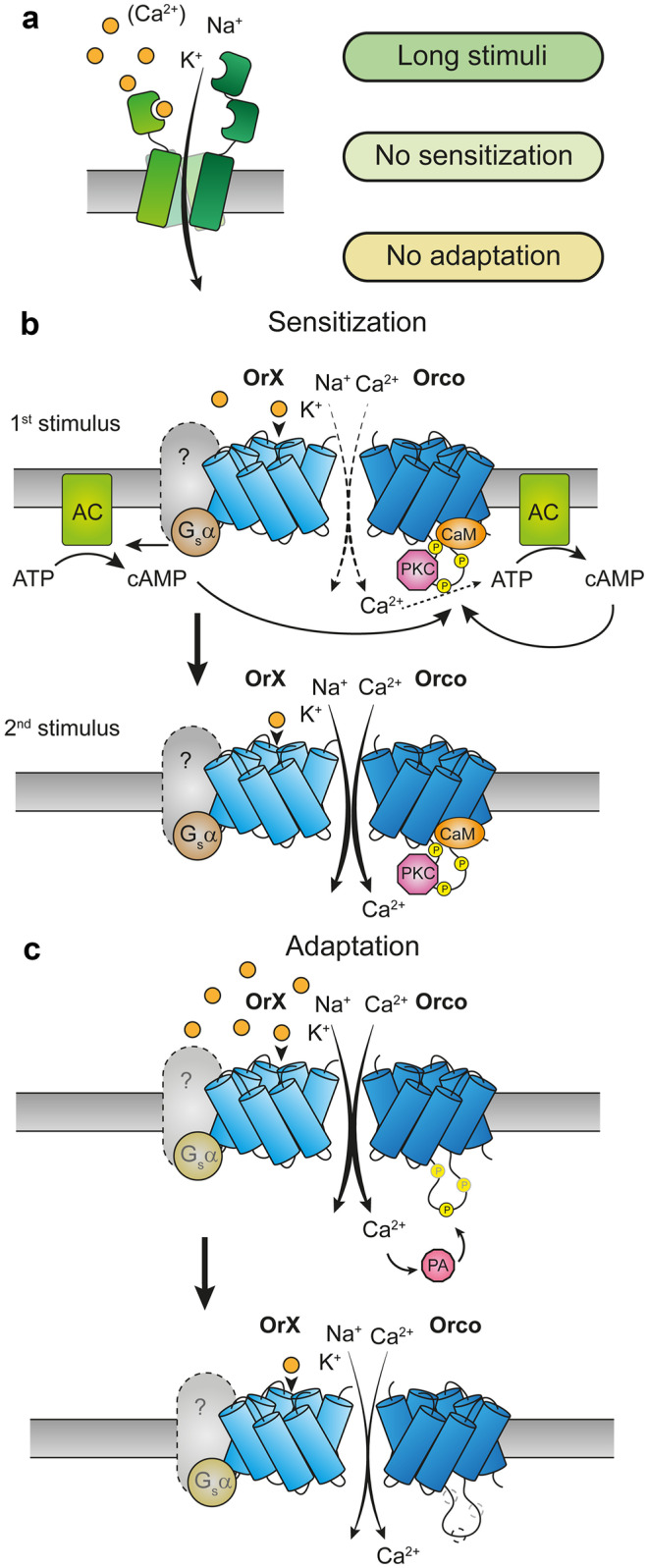
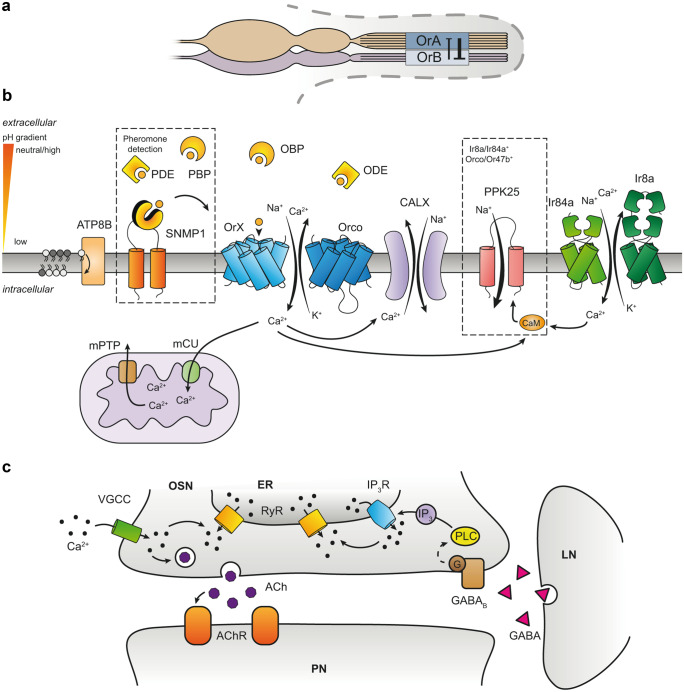
The majority of insect olfactory receptors belong to two distinct protein families, the ionotropic receptors (IRs), which are related to the ionotropic glutamate receptor family, and the odorant receptors (ORs), which evolved from the gustatory receptor family. Both receptor types assemble to heteromeric ligand-gated cation channels composed of odor-specific receptor proteins and co-receptor proteins. We here present in short the current view on evolution, function, and regulation of IRs and ORs. Special attention is given on how their functional properties can meet the environmental and ecological challenges an insect has to face.
Keywords: Adaptation; Insect olfaction; Ion channel; Ionotropic receptor; Odorant receptor; Olfactory sensory neuron; Sensitization; Signal transduction.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
Insect odorant response sensitivity is tuned by metabotropically autoregulated olfactory receptors.PLoS One. 2013;8(3):e58889. doi: 10.1371/journal.pone.0058889. Epub 2013 Mar 12. PLoS One. 2013. PMID: 23554952 Free PMC article.
-
Ancient protostome origin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction.PLoS Genet. 2010 Aug 19;6(8):e1001064. doi: 10.1371/journal.pgen.1001064. PLoS Genet. 2010. PMID: 20808886 Free PMC article.
-
Complementary function and integrated wiring of the evolutionarily distinct Drosophila olfactory subsystems.J Neurosci. 2011 Sep 21;31(38):13357-75. doi: 10.1523/JNEUROSCI.2360-11.2011. J Neurosci. 2011. PMID: 21940430 Free PMC article.
-
Ionotropic receptors (IRs): chemosensory ionotropic glutamate receptors in Drosophila and beyond.Insect Biochem Mol Biol. 2013 Sep;43(9):888-97. doi: 10.1016/j.ibmb.2013.02.007. Epub 2013 Mar 1. Insect Biochem Mol Biol. 2013. PMID: 23459169 Review.
-
Olfactory signaling in insects.Prog Mol Biol Transl Sci. 2015;130:37-54. doi: 10.1016/bs.pmbts.2014.11.002. Epub 2014 Dec 17. Prog Mol Biol Transl Sci. 2015. PMID: 25623336 Review.
Cited by
-
Far-Red Absorbing Rhodopsins, Insights From Heterodimeric Rhodopsin-Cyclases.Front Mol Biosci. 2022 Jan 21;8:806922. doi: 10.3389/fmolb.2021.806922. eCollection 2021. Front Mol Biosci. 2022. PMID: 35127823 Free PMC article. Review.
-
Task-specific odorant receptor expression in worker antennae indicates that sensory filters regulate division of labor in ants.Commun Biol. 2023 Oct 2;6(1):1004. doi: 10.1038/s42003-023-05273-4. Commun Biol. 2023. PMID: 37783732 Free PMC article.
-
Structure modelling of odorant receptor from Aedes aegypti and identification of potential repellent molecules.Comput Struct Biotechnol J. 2023 Mar 6;21:2204-2214. doi: 10.1016/j.csbj.2023.03.005. eCollection 2023. Comput Struct Biotechnol J. 2023. PMID: 37013002 Free PMC article.
-
Genome-Wide Identification and Evolutionary Analysis of Ionotropic Receptors Gene Family: Insights into Olfaction Ability Evolution and Antennal Expression Patterns in Oratosquilla oratoria.Animals (Basel). 2025 Mar 16;15(6):852. doi: 10.3390/ani15060852. Animals (Basel). 2025. PMID: 40150381 Free PMC article.
-
Lineage-Specific Class-A GPCR Dynamics Reflect Diverse Chemosensory Adaptations in Lophotrochozoa.Mol Biol Evol. 2025 Mar 5;42(3):msaf042. doi: 10.1093/molbev/msaf042. Mol Biol Evol. 2025. PMID: 39943858 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
