

Altered Membrane Mechanics Provides a Receptor-Independent Pathway for Serotonin Action
- PMID: 33502812
- PMCID: PMC8252079
- DOI: 10.1002/chem.202100328
Altered Membrane Mechanics Provides a Receptor-Independent Pathway for Serotonin Action
Abstract
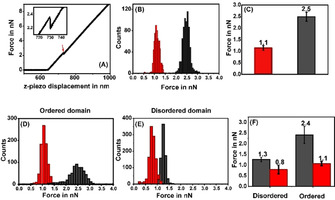
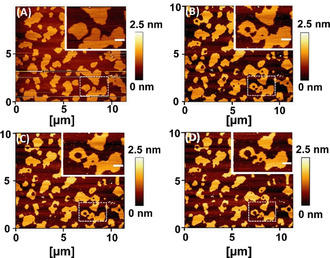
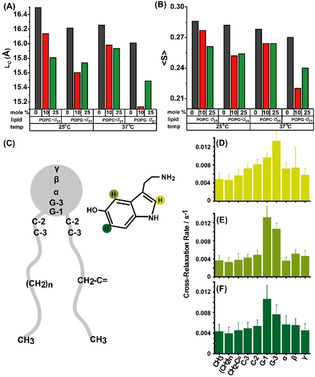
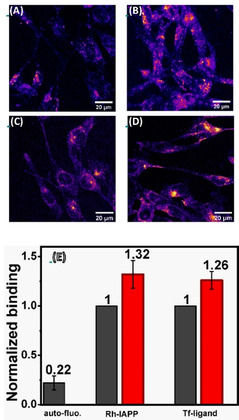
Serotonin, an important signaling molecule in humans, has an unexpectedly high lipid membrane affinity. The significance of this finding has evoked considerable speculation. Here we show that membrane binding by serotonin can directly modulate membrane properties and cellular function, providing an activity pathway completely independent of serotonin receptors. Atomic force microscopy shows that serotonin makes artificial lipid bilayers softer, and induces nucleation of liquid disordered domains inside the raft-like liquid-ordered domains. Solid-state NMR spectroscopy corroborates this data at the atomic level, revealing a homogeneous decrease in the order parameter of the lipid chains in the presence of serotonin. In the RN46A immortalized serotonergic neuronal cell line, extracellular serotonin enhances transferrin receptor endocytosis, even in the presence of broad-spectrum serotonin receptor and transporter inhibitors. Similarly, it increases the membrane binding and internalization of oligomeric peptides. Our results uncover a mode of serotonin-membrane interaction that can potentiate key cellular processes in a receptor-independent fashion.
Keywords: lipid bilayers; membrane modulation; neurotransmission; serotonin-membrane interaction; volume transmission.
© 2021 The Authors. Chemistry - A European Journal published by Wiley-VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Effects of a Serotonergic Psychedelic on the Lipid Bilayer.ACS Chem Neurosci. 2024 Nov 6;15(21):4066-4074. doi: 10.1021/acschemneuro.4c00484. Epub 2024 Oct 21. ACS Chem Neurosci. 2024. PMID: 39431923
-
Serotonergic drugs modulate the phase behavior of complex lipid bilayers.Biochimie. 2022 Dec;203:40-50. doi: 10.1016/j.biochi.2022.04.006. Epub 2022 Apr 18. Biochimie. 2022. PMID: 35447219
-
Membrane-Mediated Allosteric Action of Serotonin on a Noncognate G-Protein-Coupled Receptor.J Phys Chem Lett. 2024 Feb 15;15(6):1711-1718. doi: 10.1021/acs.jpclett.3c02340. Epub 2024 Feb 6. J Phys Chem Lett. 2024. PMID: 38319949
-
Unraveling lipid/protein interaction in model lipid bilayers by Atomic Force Microscopy.J Mol Recognit. 2011 May-Jun;24(3):387-96. doi: 10.1002/jmr.1083. J Mol Recognit. 2011. PMID: 21504015 Review.
-
Force spectroscopy as a tool to investigate the properties of supported lipid membranes.Microsc Res Tech. 2010 Oct;73(10):965-72. doi: 10.1002/jemt.20834. Microsc Res Tech. 2010. PMID: 20232466 Review.
Cited by
-
Cholesterol-Dependent Serotonin Insertion Controlled by Gangliosides in Model Lipid Membranes.Int J Mol Sci. 2024 Sep 23;25(18):10194. doi: 10.3390/ijms251810194. Int J Mol Sci. 2024. PMID: 39337677 Free PMC article.
-
Interaction of the pitavastatin with model membranes.Biochem Biophys Rep. 2021 Sep 29;28:101143. doi: 10.1016/j.bbrep.2021.101143. eCollection 2021 Dec. Biochem Biophys Rep. 2021. PMID: 34632116 Free PMC article.
-
Serotonin Signaling through Lipid Membranes.ACS Chem Neurosci. 2024 Apr 3;15(7):1298-1320. doi: 10.1021/acschemneuro.3c00823. Epub 2024 Mar 18. ACS Chem Neurosci. 2024. PMID: 38499042 Free PMC article. Review.
-
Selective Advantages of Synapses in Evolution.Front Cell Dev Biol. 2021 Aug 20;9:726563. doi: 10.3389/fcell.2021.726563. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34490275 Free PMC article. No abstract available.
-
Mechanistic Understanding from Molecular Dynamics in Pharmaceutical Research 2: Lipid Membrane in Drug Design.Pharmaceuticals (Basel). 2021 Oct 19;14(10):1062. doi: 10.3390/ph14101062. Pharmaceuticals (Basel). 2021. PMID: 34681286 Free PMC article. Review.
References
-
- Kandel E. R., Jessell T. M., Siegelbaum S., Principles of Neural Science McGraw-Hill, New York, 2021.
-
- Cantor R. S., Biochemistry 2003, 42, 11891–11897. - PubMed
-
- Revill A. L., Chu N. Y., Ma L., LeBlancq M. J., Dickson C. T., Funk G. D., J. Physiol. 2019, 597, 3183–3201. - PubMed
-
- Saitow F., Murano M., Suzuki H., J. Neurophysiol. 2009, 101, 1361–1374. - PubMed
-
- Murano M., Saitow F., Suzuki H., Neuroscience 2011, 172, 118–128. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
