

Magnesium Signaling in Plants
- PMID: 33503839
- PMCID: PMC7865908
- DOI: 10.3390/ijms22031159
Magnesium Signaling in Plants
Abstract
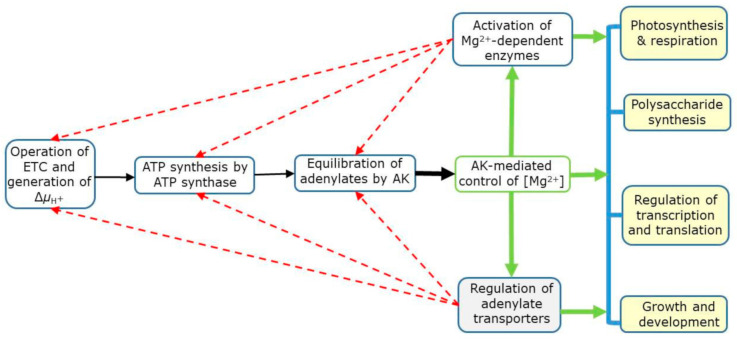
Free magnesium (Mg2+) is a signal of the adenylate (ATP+ADP+AMP) status in the cells. It results from the equilibrium of adenylate kinase (AK), which uses Mg-chelated and Mg-free adenylates as substrates in both directions of its reaction. The AK-mediated primary control of intracellular [Mg2+] is finely interwoven with the operation of membrane-bound adenylate- and Mg2+-translocators, which in a given compartment control the supply of free adenylates and Mg2+ for the AK-mediated equilibration. As a result, [Mg2+] itself varies both between and within the compartments, depending on their energetic status and environmental clues. Other key nucleotide-utilizing/producing enzymes (e.g., nucleoside diphosphate kinase) may also be involved in fine-tuning of the intracellular [Mg2+]. Changes in [Mg2+] regulate activities of myriads of Mg-utilizing/requiring enzymes, affecting metabolism under both normal and stress conditions, and impacting photosynthetic performance, respiration, phloem loading and other processes. In compartments controlled by AK equilibrium (cytosol, chloroplasts, mitochondria, nucleus), the intracellular [Mg2+] can be calculated from total adenylate contents, based on the dependence of the apparent equilibrium constant of AK on [Mg2+]. Magnesium signaling, reflecting cellular adenylate status, is likely widespread in all eukaryotic and prokaryotic organisms, due simply to the omnipresent nature of AK and to its involvement in adenylate equilibration.
Keywords: adenylate energy charge; adenylate kinase; cellular magnesium; free magnesium; nucleoside diphosphate kinase; thermodynamic buffering.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Magnesium and cell energetics: At the junction of metabolism of adenylate and non-adenylate nucleotides.J Plant Physiol. 2023 Jan;280:153901. doi: 10.1016/j.jplph.2022.153901. Epub 2022 Dec 12. J Plant Physiol. 2023. PMID: 36549033 Review.
-
Membrane potential, adenylate levels and Mg2+ are interconnected via adenylate kinase equilibrium in plant cells.Biochim Biophys Acta. 2003 Dec 8;1607(2-3):111-9. doi: 10.1016/j.bbabio.2003.09.005. Biochim Biophys Acta. 2003. PMID: 14670601 Review.
-
Implications of adenylate kinase-governed equilibrium of adenylates on contents of free magnesium in plant cells and compartments.Biochem J. 2001 Nov 15;360(Pt 1):225-31. doi: 10.1042/0264-6021:3600225. Biochem J. 2001. PMID: 11696011 Free PMC article.
-
Equilibration of adenylates in the mitochondrial intermembrane space maintains respiration and regulates cytosolic metabolism.J Exp Bot. 2006;57(10):2133-41. doi: 10.1093/jxb/erl006. Epub 2006 Jun 23. J Exp Bot. 2006. PMID: 16798851 Review.
-
Optimization of ATP synthase function in mitochondria and chloroplasts via the adenylate kinase equilibrium.Front Plant Sci. 2015 Jan 28;6:10. doi: 10.3389/fpls.2015.00010. eCollection 2015. Front Plant Sci. 2015. PMID: 25674099 Free PMC article.
Cited by
-
Exploring Redox Modulation of Plant UDP-Glucose Pyrophosphorylase.Int J Mol Sci. 2023 May 17;24(10):8914. doi: 10.3390/ijms24108914. Int J Mol Sci. 2023. PMID: 37240260 Free PMC article.
-
Infection-relevant conditions dictate differential versus coordinate expression of Salmonella chaperones and cochaperones.mBio. 2025 May 14;16(5):e0022725. doi: 10.1128/mbio.00227-25. Epub 2025 Mar 31. mBio. 2025. PMID: 40162747 Free PMC article.
-
The Role of Transmembrane Proteins in Plant Growth, Development, and Stress Responses.Int J Mol Sci. 2022 Nov 7;23(21):13627. doi: 10.3390/ijms232113627. Int J Mol Sci. 2022. PMID: 36362412 Free PMC article. Review.
-
Artemisia and the Elements: A Botanical Symphony of Minerals and Metals.Biol Trace Elem Res. 2025 Jun 20. doi: 10.1007/s12011-025-04706-x. Online ahead of print. Biol Trace Elem Res. 2025. PMID: 40540239 Review.
-
Effects of Magnesium on Transcriptome and Physicochemical Index of Tea Leaves.Plants (Basel). 2023 Apr 28;12(9):1810. doi: 10.3390/plants12091810. Plants (Basel). 2023. PMID: 37176867 Free PMC article.
References
-
- Dorenstouter H., Pieters G., Findenegg G. Distribution of magnesium between chlorophyll and other photosynthetic functions in magnesium deficient “sun”; and “shade”; leaves of poplar. J. Plant Nutr. 1985;8:1089–1101. doi: 10.1080/01904168509363409. - DOI
-
- Martin M.H., Marschner H. The mineral nutrition of higher plants. J. Ecol. 1988;76:1250. doi: 10.2307/2260650. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
