

Diversity and versatility of p38 kinase signalling in health and disease
- PMID: 33504982
- PMCID: PMC7838852
- DOI: 10.1038/s41580-020-00322-w
Diversity and versatility of p38 kinase signalling in health and disease
Abstract
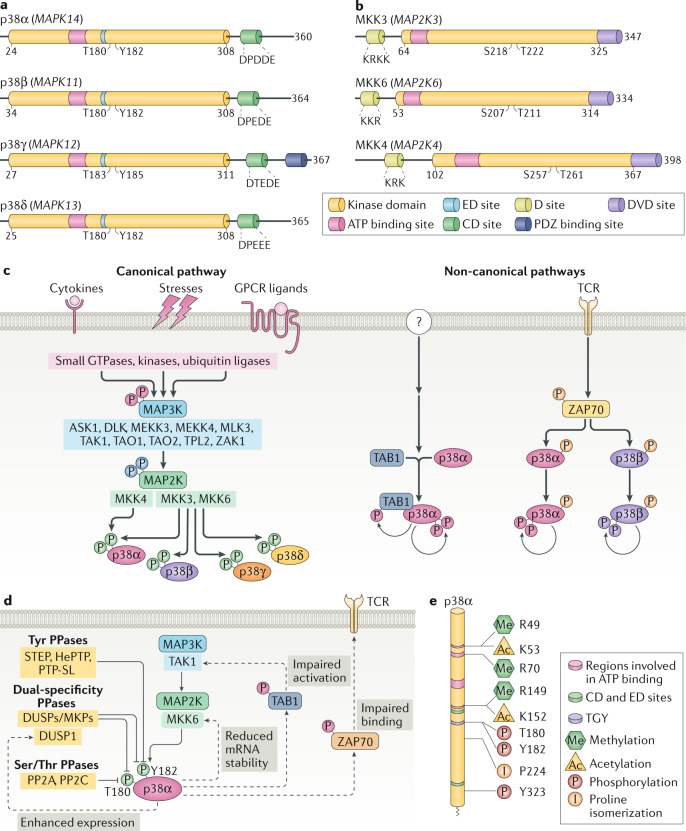
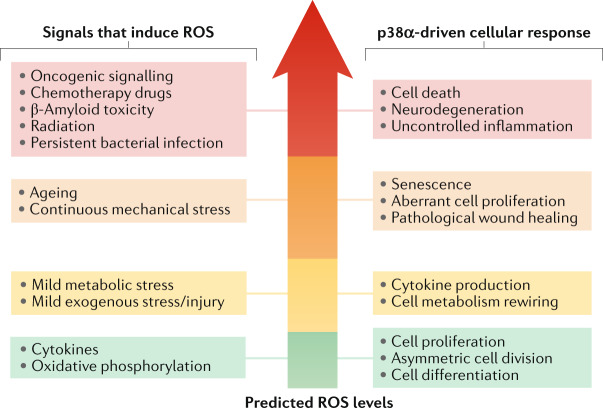
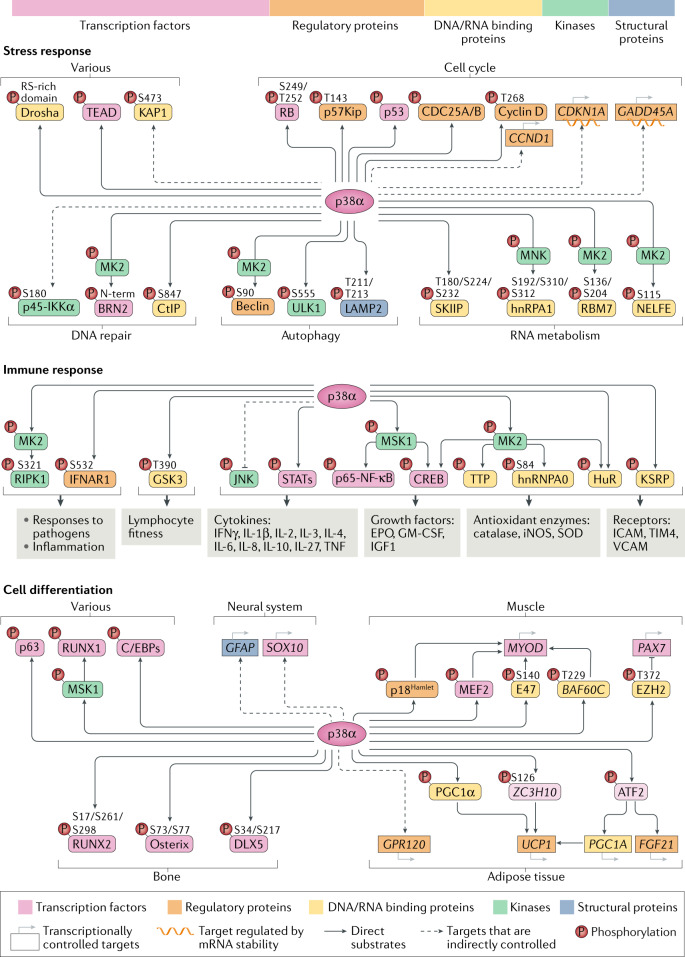
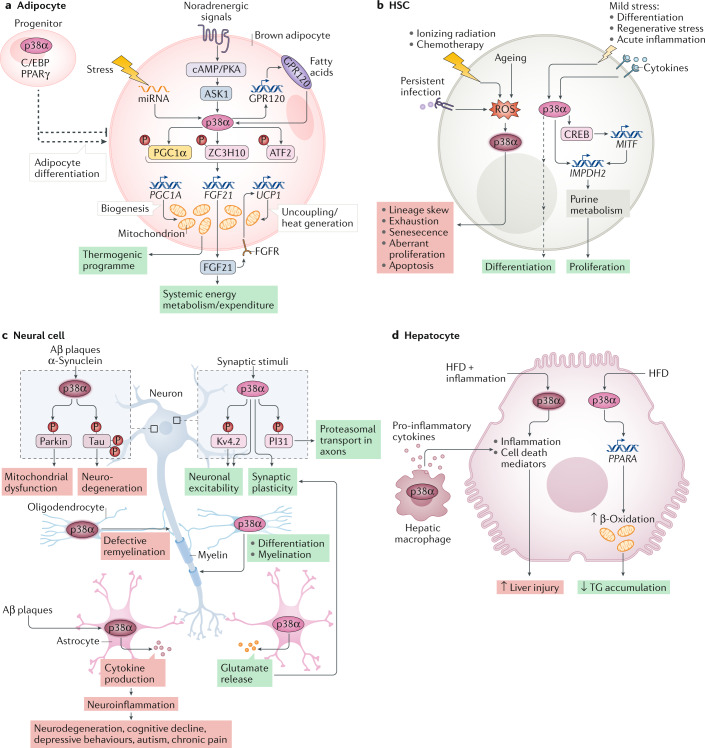
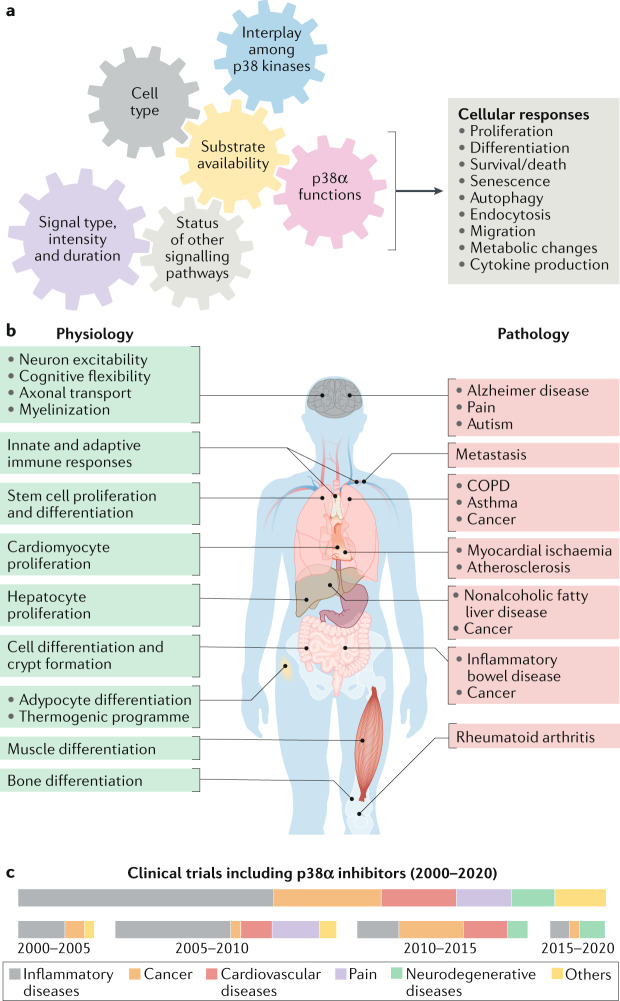
The ability of cells to deal with different types of stressful situations in a precise and coordinated manner is key for survival and involves various signalling networks. Over the past 25 years, p38 kinases - in particular, p38α - have been implicated in the cellular response to stress at many levels. These span from environmental and intracellular stresses, such as hyperosmolarity, oxidative stress or DNA damage, to physiological situations that involve important cellular changes such as differentiation. Given that p38α controls a plethora of functions, dysregulation of this pathway has been linked to diseases such as inflammation, immune disorders or cancer, suggesting the possibility that targeting p38α could be of therapeutic interest. In this Review, we discuss the organization of this signalling pathway focusing on the diversity of p38α substrates, their mechanisms and their links to particular cellular functions. We then address how the different cellular responses can be generated depending on the signal received and the cell type, and highlight the roles of this kinase in human physiology and in pathological contexts.
Conflict of interest statement
IRB Barcelona and ICREA have filed a patent application on compounds that modulate p38α autophosphorylation (WO2020120576). A.R.N. is a named inventor on this application. B.C. declares no competing interests.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
