

Translational Control by Ribosome Pausing in Bacteria: How a Non-uniform Pace of Translation Affects Protein Production and Folding
- PMID: 33505387
- PMCID: PMC7829197
- DOI: 10.3389/fmicb.2020.619430
Translational Control by Ribosome Pausing in Bacteria: How a Non-uniform Pace of Translation Affects Protein Production and Folding
Abstract
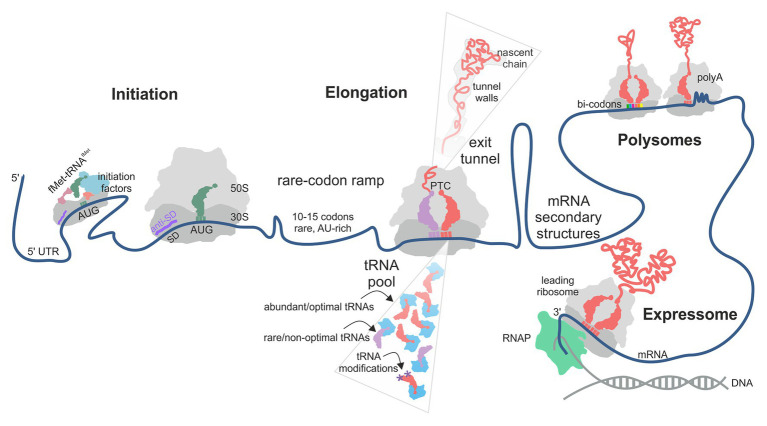
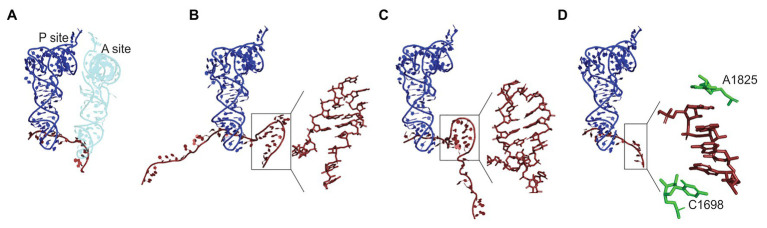
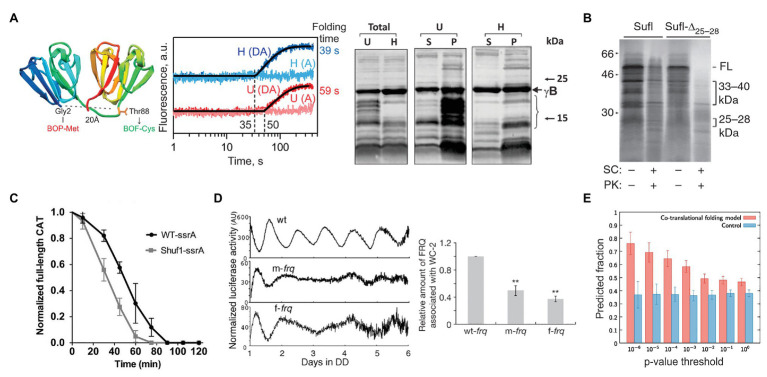
Protein homeostasis of bacterial cells is maintained by coordinated processes of protein production, folding, and degradation. Translational efficiency of a given mRNA depends on how often the ribosomes initiate synthesis of a new polypeptide and how quickly they read the coding sequence to produce a full-length protein. The pace of ribosomes along the mRNA is not uniform: periods of rapid synthesis are separated by pauses. Here, we summarize recent evidence on how ribosome pausing affects translational efficiency and protein folding. We discuss the factors that slow down translation elongation and affect the quality of the newly synthesized protein. Ribosome pausing emerges as important factor contributing to the regulatory programs that ensure the quality of the proteome and integrate the cellular and environmental cues into regulatory circuits of the cell.
Keywords: cotranslational folding; nascent peptide; prokaryotes; ribosome pausing; tRNA; translation; translation efficiency.
Copyright © 2021 Samatova, Daberger, Liutkute and Rodnina.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources