

Cryo-EM structure of human mitochondrial HSPD1
- PMID: 33506187
- PMCID: PMC7814154
- DOI: 10.1016/j.isci.2020.102022
Cryo-EM structure of human mitochondrial HSPD1
Abstract
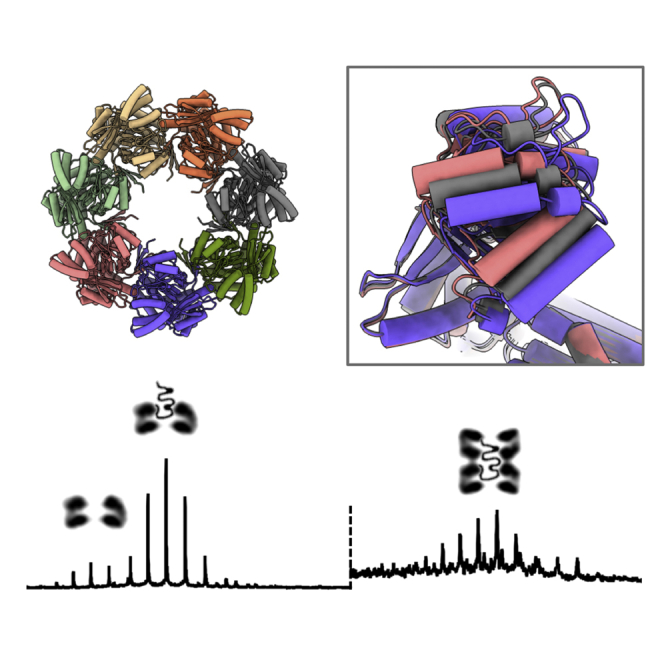
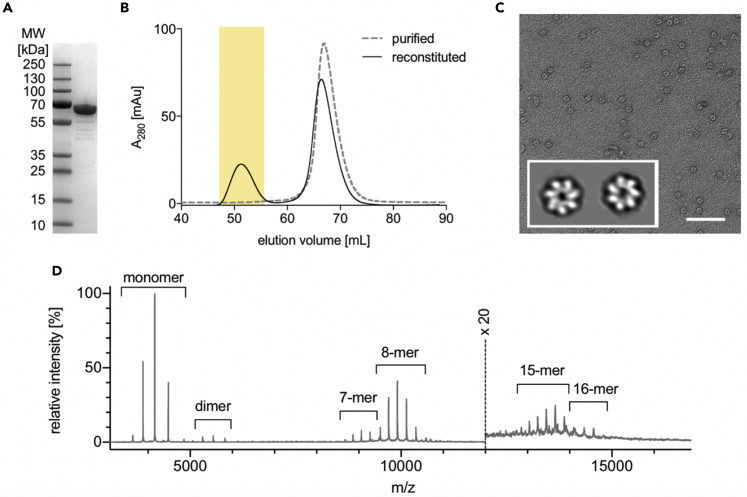
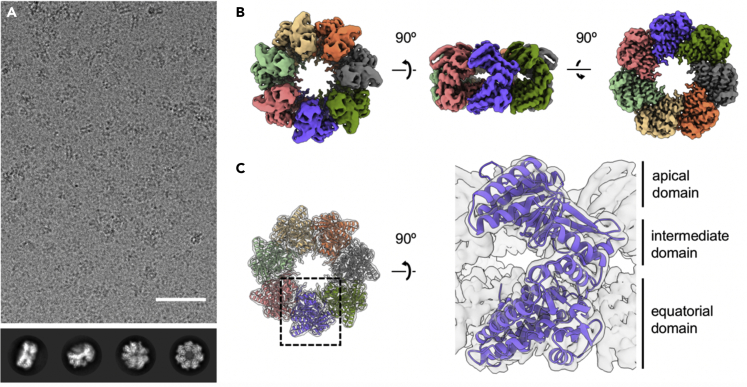
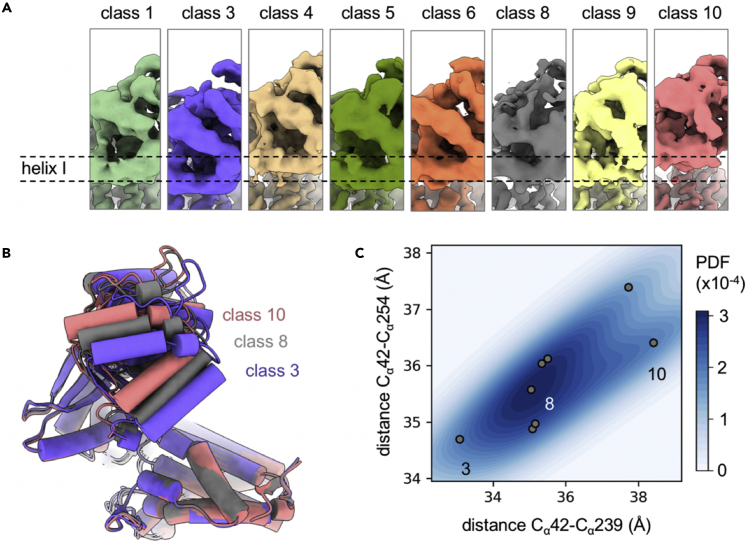
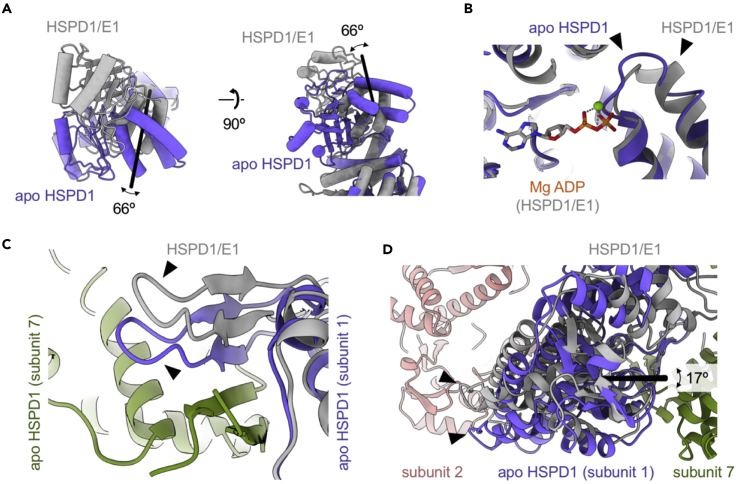
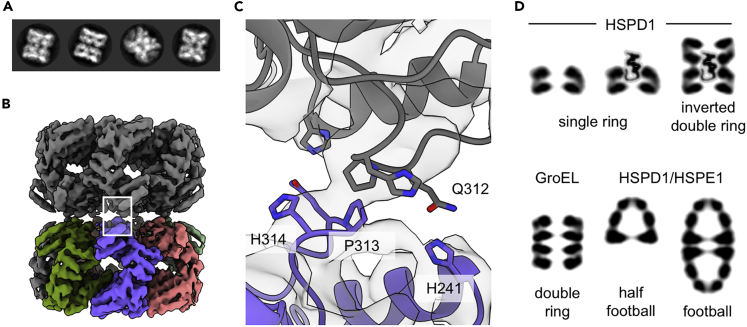
Chaperonins play an important role in folding newly synthesized or translocated proteins in all organisms. The bacterial chaperonin GroEL has served as a model system for the understanding of these proteins. In comparison, its human homolog, known as mitochondrial heat shock protein family member D1 (HSPD1) is poorly understood. Here, we present the structure of HSPD1 in the apo state determined by cryo-electron microscopy (cryo-EM). Unlike GroEL, HSPD1 forms mostly single ring assemblies in the absence of co-chaperonin (HSPE1). Comparison with GroEL shows a rotation and increased flexibility of the apical domain. Together with published structures of the HSPD1/HSPE1 co-chaperonin complex, this work gives insight into the structural changes that occur during the catalytic cycle. This new understanding of HSPD1 structure and its rearrangements upon complex formation may provide new insights for the development of HSPD1-targeting treatments against a diverse range of diseases including glioblastoma.
Keywords: Molecular Biology; Molecular Structure.
© 2020.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Bartolucci C., Lamba D., Grazulis S., Manakova E., Heumann H. Crystal structure of wild-type chaperonin GroEL. J. Mol. Biol. 2005;354:940–951. - PubMed
-
- Braig K., Otwinowski Z., Hegde R., Boisvert D.C., Joachimiak A., Horwich A.L., Sigler P.B. The crystal structure of the bacterial chaperonin GroEL at 2.8 A. Nature. 1994;371:578–586. - PubMed
-
- Bukau B., Horwich A.L. The Hsp70 and Hsp60 chaperone machines. Cell. 1998;92:351–366. - PubMed
-
- Cappello F., Marino Gammazza A., Palumbo Piccionello A., Campanella C., Pace A., Conway de Macario E., Macario A.J. Hsp60 chaperonopathies and chaperonotherapy: targets and agents. Expert Opin. Ther. Targets. 2014;18:185–208. - PubMed
-
- Chen L., Sigler P.B. The crystal structure of a GroEL/peptide complex: plasticity as a basis for substrate diversity. Cell. 1999;99:757–768. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
