

Avoiding Drug Resistance in HIV Reverse Transcriptase
- PMID: 33507067
- PMCID: PMC8149104
- DOI: 10.1021/acs.chemrev.0c00967
Avoiding Drug Resistance in HIV Reverse Transcriptase
Abstract
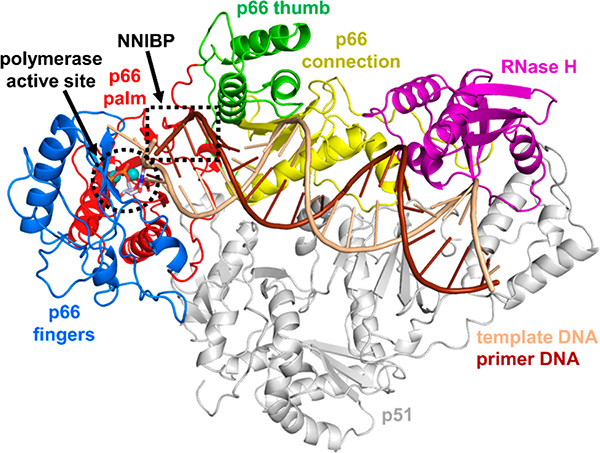
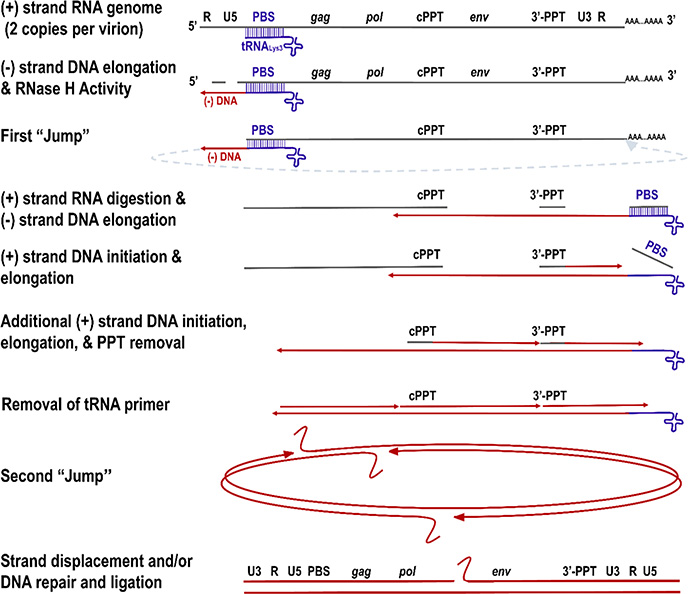
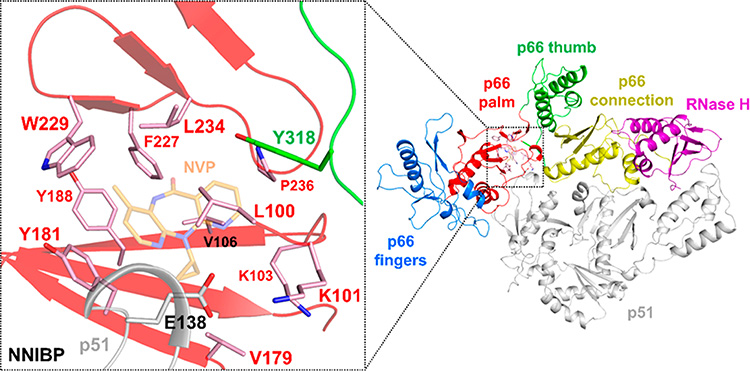
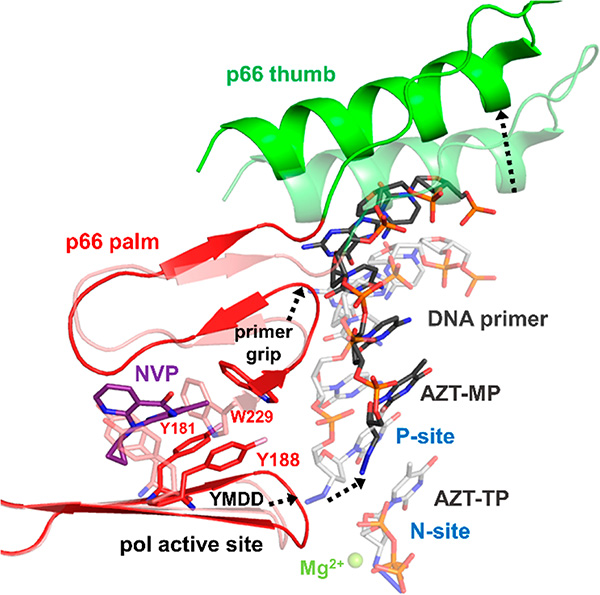
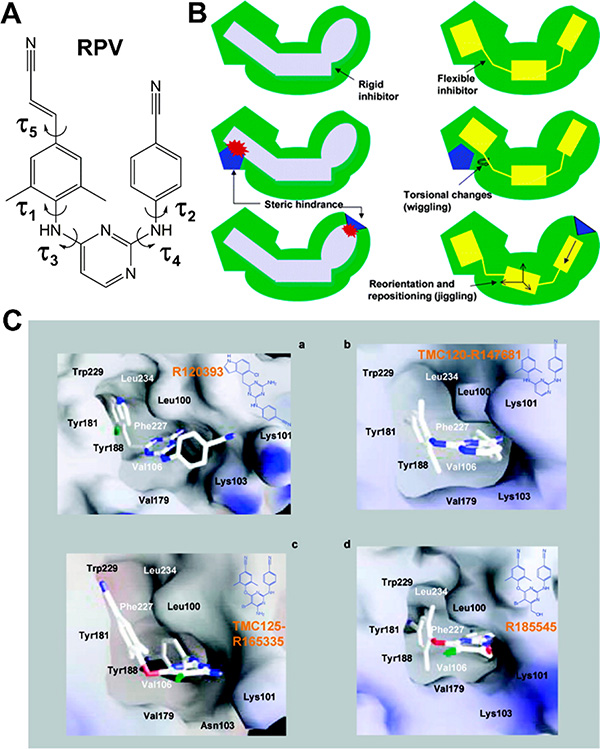
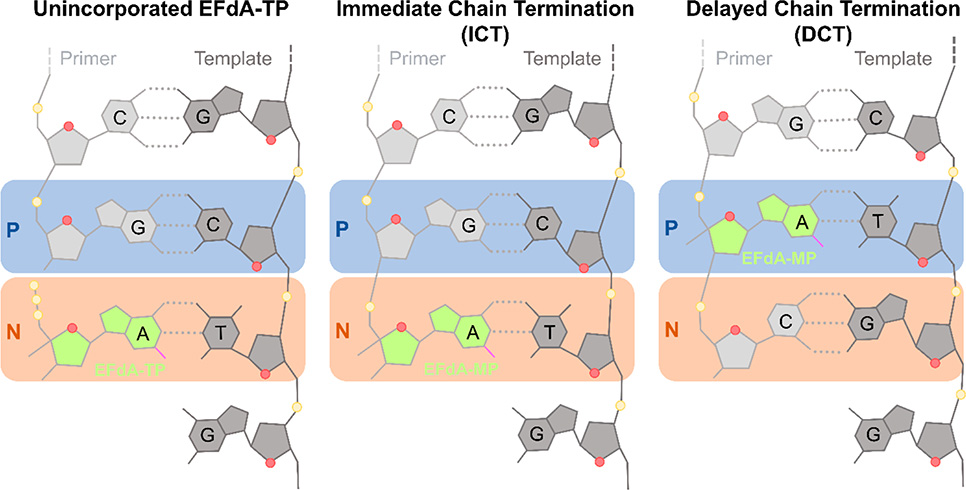
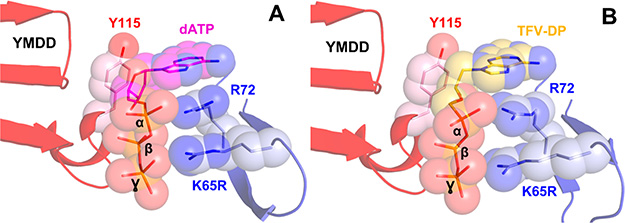
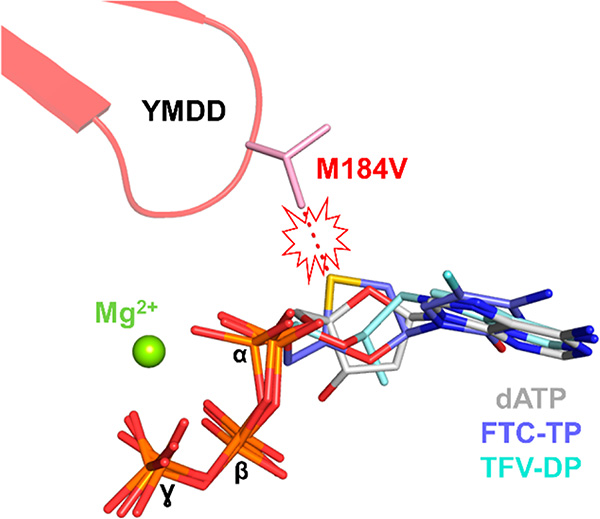
HIV reverse transcriptase (RT) is an enzyme that plays a major role in the replication cycle of HIV and has been a key target of anti-HIV drug development efforts. Because of the high genetic diversity of the virus, mutations in RT can impart resistance to various RT inhibitors. As the prevalence of drug resistance mutations is on the rise, it is necessary to design strategies that will lead to drugs less susceptible to resistance. Here we provide an in-depth review of HIV reverse transcriptase, current RT inhibitors, novel RT inhibitors, and mechanisms of drug resistance. We also present novel strategies that can be useful to overcome RT's ability to escape therapies through drug resistance. While resistance may not be completely avoidable, designing drugs based on the strategies and principles discussed in this review could decrease the prevalence of drug resistance.
Conflict of interest statement
The authors declare no competing financial interest.
Figures





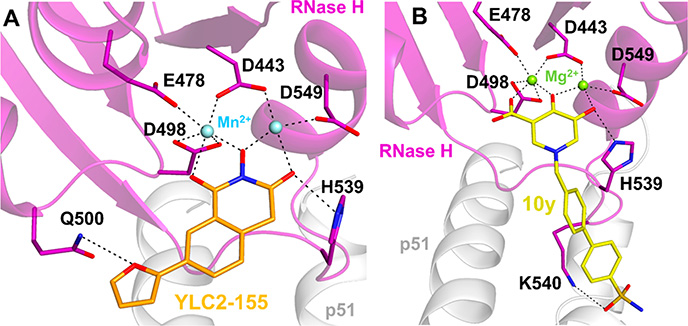

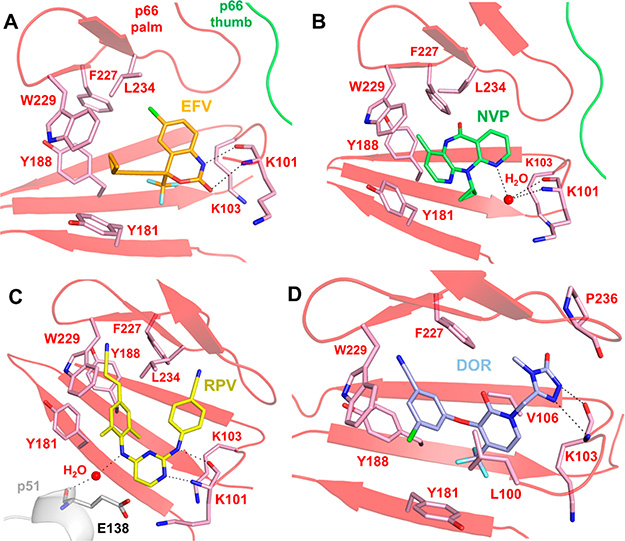

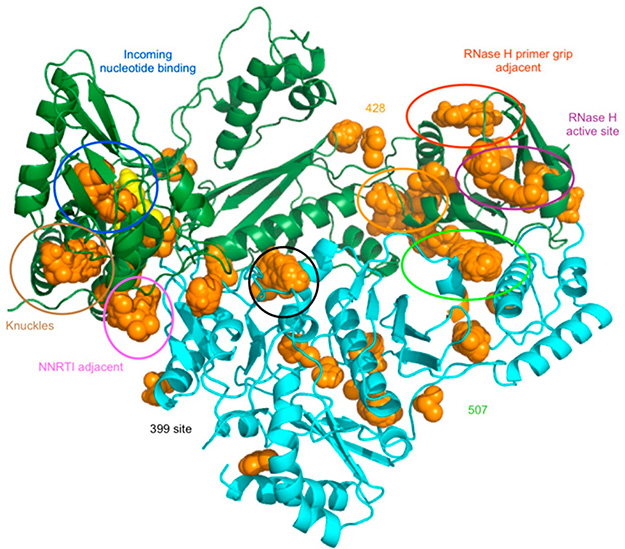
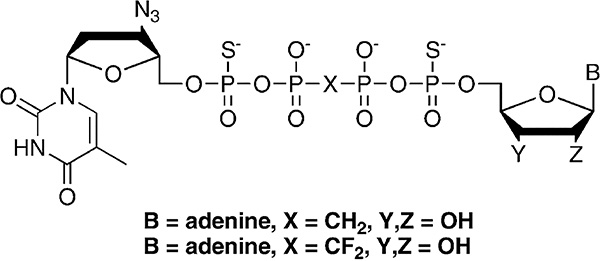
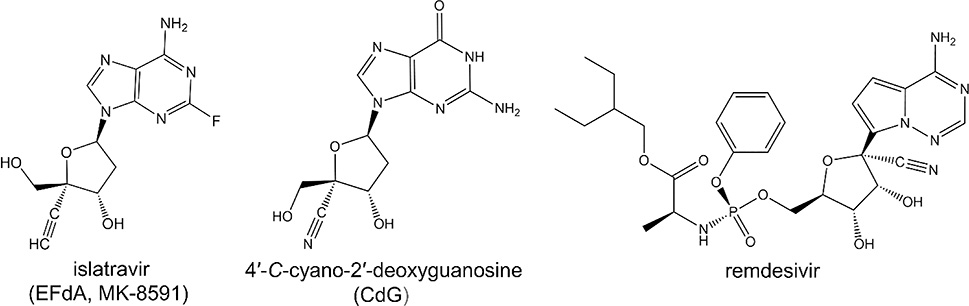
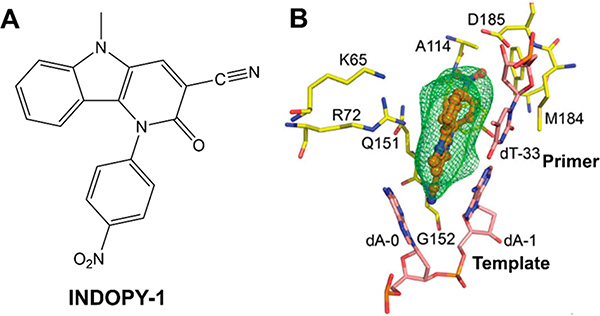
References
-
- Lucas S; Nelson AM HIV and the spectrum of human disease. J. Pathol. 2015, 235, 229–241. - PubMed
-
- Shafer RW; Vuitton DA Highly active antiretroviral therapy (HAART) for the treatment of infection with human immunodeficiency virus type 1. Biomed. Pharmacother. 1999, 53, 73–86. - PubMed
-
- Eggleton JS; Nagalli S Highly Active Antiretroviral Therapy (HAART); StatPearls: Treasure Island, FL, 2020. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
