

Allele-specific editing ameliorates dominant retinitis pigmentosa in a transgenic mouse model
- PMID: 33508235
- PMCID: PMC7896132
- DOI: 10.1016/j.ajhg.2021.01.006
Allele-specific editing ameliorates dominant retinitis pigmentosa in a transgenic mouse model
Abstract
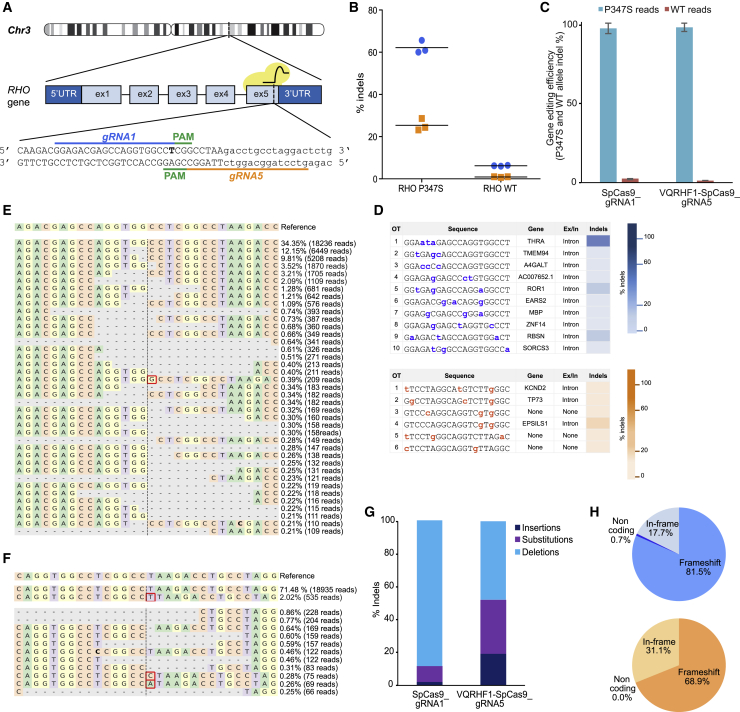
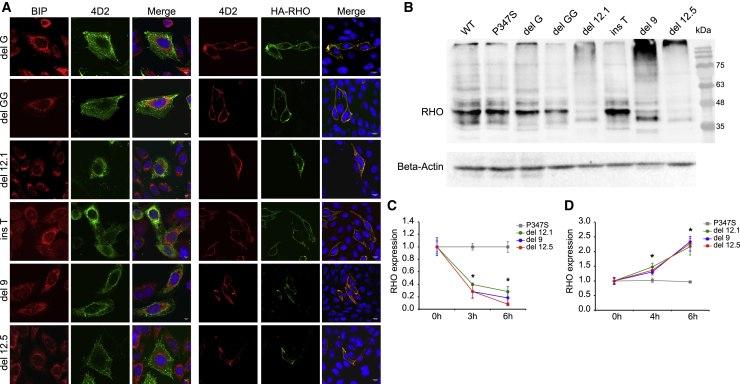
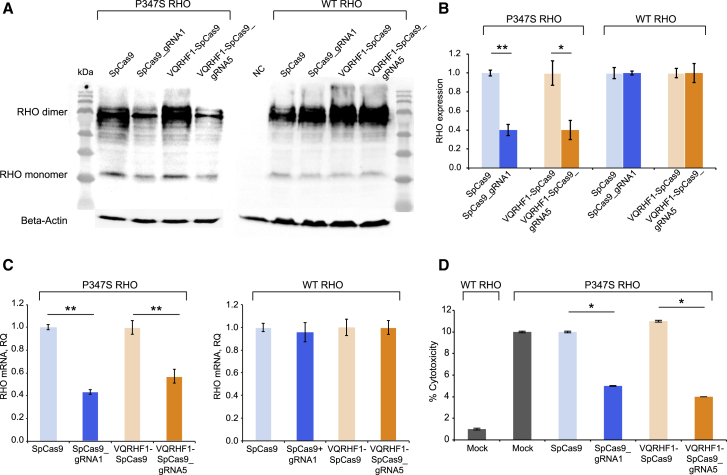
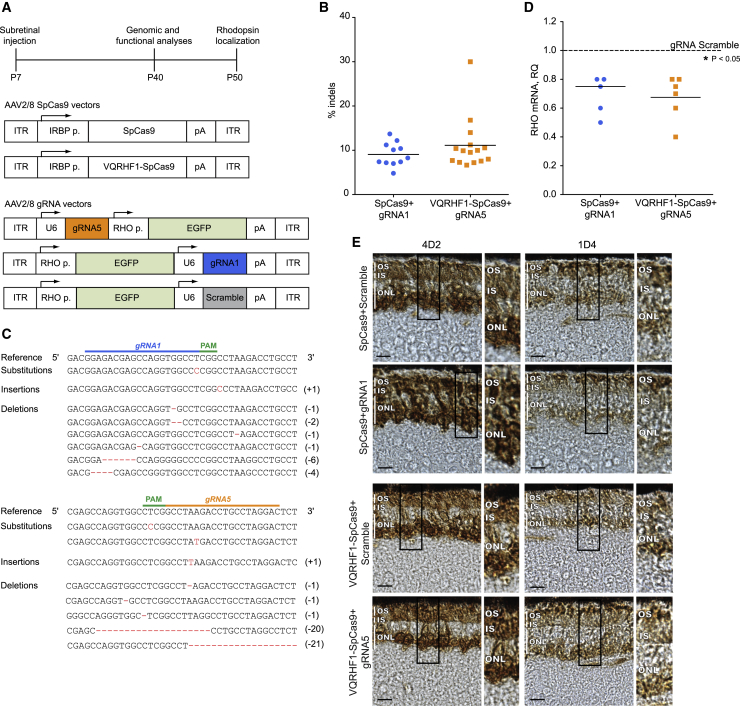
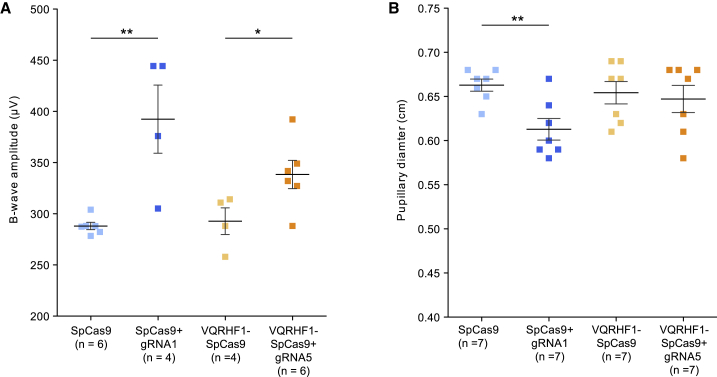
Retinitis pigmentosa (RP) is a group of progressive retinal degenerations of mostly monogenic inheritance, which cause blindness in about 1:3,500 individuals worldwide. Heterozygous variants in the rhodopsin (RHO) gene are the most common cause of autosomal dominant RP (adRP). Among these, missense variants at C-terminal proline 347, such as p.Pro347Ser, cause severe adRP recurrently in European affected individuals. Here, for the first time, we use CRISPR/Cas9 to selectively target the p.Pro347Ser variant while preserving the wild-type RHO allele in vitro and in a mouse model of adRP. Detailed in vitro, genomic, and biochemical characterization of the rhodopsin C-terminal editing demonstrates a safe downregulation of p.Pro347Ser expression leading to partial recovery of photoreceptor function in a transgenic mouse model treated with adeno-associated viral vectors. This study supports the safety and efficacy of CRISPR/Cas9-mediated allele-specific editing and paves the way for a permanent and precise correction of heterozygous variants in dominantly inherited retinal diseases.
Keywords: AAV vector; CRISPR-Cas9 editing; Rhodopsin; retinitis pigmentosa; transgenic mice.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
C.P., M.L., D.B., A.A., and A.R. are listed as inventors on a patent application related to this work.
Figures
References
-
- Verbakel S.K., van Huet R.A.C., Boon C.J.F., den Hollander A.I., Collin R.W.J., Klaver C.C.W., Hoyng C.B., Roepman R., Klevering B.J. Non-syndromic retinitis pigmentosa. Prog. Retin. Eye Res. 2018;66:157–186. - PubMed
-
- Benati D., Patrizi C., Recchia A. Gene editing prospects for treating inherited retinal diseases. J. Med. Genet. 2020;57:437–444. - PubMed
-
- Dias M.F., Joo K., Kemp J.A., Fialho S.L., da Silva Cunha A., Jr., Woo S.J., Kwon Y.J. Molecular genetics and emerging therapies for retinitis pigmentosa: Basic research and clinical perspectives. Prog. Retin. Eye Res. 2018;63:107–131. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
