

Mispair-bound human MutS-MutL complex triggers DNA incisions and activates mismatch repair
- PMID: 33510387
- PMCID: PMC8089094
- DOI: 10.1038/s41422-021-00468-y
Mispair-bound human MutS-MutL complex triggers DNA incisions and activates mismatch repair
Abstract
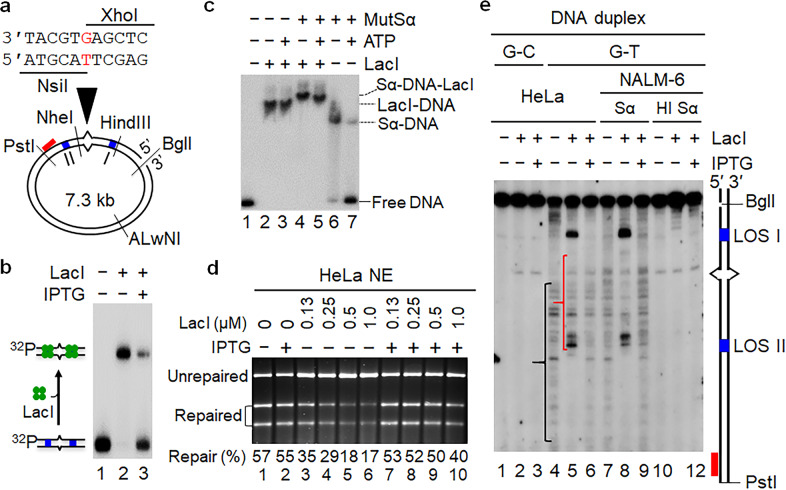
DNA mismatch repair (MMR) relies on MutS and MutL ATPases for mismatch recognition and strand-specific nuclease recruitment to remove mispaired bases in daughter strands. However, whether the MutS-MutL complex coordinates MMR by ATP-dependent sliding on DNA or protein-protein interactions between the mismatch and strand discrimination signal is ambiguous. Using functional MMR assays and systems preventing proteins from sliding, we show that sliding of human MutSα is required not for MMR initiation, but for final mismatch removal. MutSα recruits MutLα to form a mismatch-bound complex, which initiates MMR by nicking the daughter strand 5' to the mismatch. Exonuclease 1 (Exo1) is then recruited to the nick and conducts 5' → 3' excision. ATP-dependent MutSα dissociation from the mismatch is necessary for Exo1 to remove the mispaired base when the excision reaches the mismatch. Therefore, our study has resolved a long-standing puzzle, and provided new insights into the mechanism of MMR initiation and mispair removal.
Conflict of interest statement
The authors declare no competing interests.
Figures






References
-
- Modrich P, Lahue R. Mismatch repair in replication fidelity, genetic recombination, and cancer biology. Annu. Rev. Biochem. 1996;65:101–133. - PubMed
-
- Kolodner RD, Marsischky GT. Eukaryotic DNA mismatch repair. Curr. Opin. Genet. Dev. 1999;9:89–96. - PubMed
-
- Kunkel TA, Erie DA. DNA mismatch repair. Annu. Rev. Biochem. 2005;74:681–710. - PubMed
-
- Li GM. Mechanisms and functions of DNA mismatch repair. Cell Res. 2008;18:85–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
