

Activated protein C has a regulatory role in factor VIII function
- PMID: 33512448
- PMCID: PMC8109017
- DOI: 10.1182/blood.2020007562
Activated protein C has a regulatory role in factor VIII function
Abstract
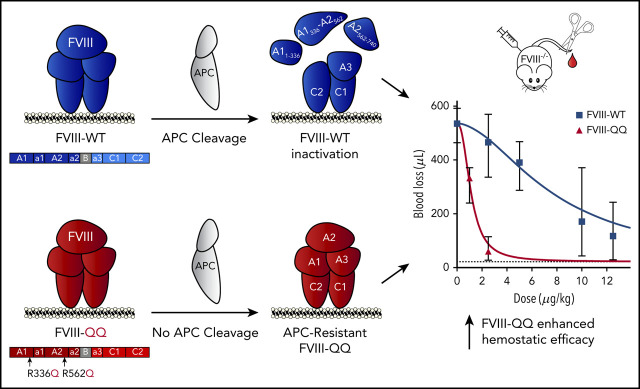
Mechanisms thought to regulate activated factor VIII (FVIIIa) cofactor function include A2-domain dissociation and activated protein C (APC) cleavage. Unlike A2-domain dissociation, there is no known phenotype associated with altered APC cleavage of FVIII, and biochemical studies have suggested APC plays a marginal role in FVIIIa regulation. However, the in vivo contribution of FVIIIa inactivation by APC is unexplored. Here we compared wild-type B-domainless FVIII (FVIII-WT) recombinant protein with an APC-resistant FVIII variant (FVIII-R336Q/R562Q; FVIII-QQ). FVIII-QQ demonstrated expected APC resistance without other changes in procoagulant function or A2-domain dissociation. In plasma-based studies, FVIII-WT/FVIIIa-WT demonstrated dose-dependent sensitivity to APC with or without protein S, whereas FVIII-QQ/FVIIIa-QQ did not. Importantly, FVIII-QQ demonstrated approximately fivefold increased procoagulant function relative to FVIII-WT in the tail clip and ferric chloride injury models in hemophilia A (HA) mice. To minimize the contribution of FV inactivation by APC in vivo, a tail clip assay was performed in homozygous HA/FV Leiden (FVL) mice infused with FVIII-QQ or FVIII-WT in the presence or absence of monoclonal antibody 1609, an antibody that blocks murine PC/APC hemostatic function. FVIII-QQ again demonstrated enhanced hemostatic function in HA/FVL mice; however, FVIII-QQ and FVIII-WT performed analogously in the presence of the PC/APC inhibitory antibody, indicating the increased hemostatic effect of FVIII-QQ was APC specific. Our data demonstrate APC contributes to the in vivo regulation of FVIIIa, which has the potential to be exploited to develop novel HA therapeutics.
© 2021 by The American Society of Hematology.
Conflict of interest statement
Conflict-of-interest disclosure: R.M.C. and L.A.G. hold intellectual property related to this work. The remaining authors declare no competing financial interests.
Figures






Similar articles
-
Factor IXa and Factor X Influence Factor VIIIa Stability and Inactivation Mechanisms In Vitro and In Vivo.Blood. 2025 Jul 15:blood.2024027705. doi: 10.1182/blood.2024027705. Online ahead of print. Blood. 2025. PMID: 40663775
-
Contribution of factor VIII light-chain residues 2007-2016 to an activated protein C-interactive site.Thromb Haemost. 2013 Feb;109(2):187-98. doi: 10.1160/TH12-08-0561. Epub 2012 Dec 6. Thromb Haemost. 2013. PMID: 23224054 Free PMC article.
-
Intrinsic stability and functional properties of disulfide bond-stabilized coagulation factor VIIIa variants.J Thromb Haemost. 2006 Jun;4(6):1315-22. doi: 10.1111/j.1538-7836.2006.01951.x. J Thromb Haemost. 2006. PMID: 16706977
-
Activated protein C resistance and thrombosis: molecular mechanisms of hypercoagulable state due to FVR506Q mutation.Semin Thromb Hemost. 1999;25(3):273-89. doi: 10.1055/s-2007-994931. Semin Thromb Hemost. 1999. PMID: 10443959 Review.
-
Laboratory Monitoring in Emicizumab-Treated Persons with Hemophilia A.Thromb Haemost. 2019 Sep;119(9):1384-1393. doi: 10.1055/s-0039-1692427. Epub 2019 Jun 16. Thromb Haemost. 2019. PMID: 31203578 Review.
Cited by
-
Multiyear Factor VIII Expression after AAV Gene Transfer for Hemophilia A.N Engl J Med. 2021 Nov 18;385(21):1961-1973. doi: 10.1056/NEJMoa2104205. N Engl J Med. 2021. PMID: 34788507 Free PMC article. Clinical Trial.
-
Roctavian gene therapy for hemophilia A.Blood Adv. 2024 Oct 8;8(19):5179-5189. doi: 10.1182/bloodadvances.2023011847. Blood Adv. 2024. PMID: 38991118 Free PMC article. Review.
-
Factor VIII trafficking to CD4+ T cells shapes its immunogenicity and requires several types of antigen-presenting cells.Blood. 2023 Jul 20;142(3):290-305. doi: 10.1182/blood.2022018937. Blood. 2023. PMID: 37192286 Free PMC article.
-
Factor IXa variants resistant to plasma inhibitors enhance clot formation in vivo.Blood. 2023 Apr 20;141(16):2022-2032. doi: 10.1182/blood.2022018083. Blood. 2023. PMID: 36724452 Free PMC article.
-
Adeno-Associated Virus Gene Therapy for Hemophilia.Annu Rev Med. 2023 Jan 27;74:231-247. doi: 10.1146/annurev-med-043021-033013. Epub 2022 Sep 14. Annu Rev Med. 2023. PMID: 36103998 Free PMC article. Review.
References
-
- Eaton D, Rodriguez H, Vehar GA. Proteolytic processing of human factor VIII. Correlation of specific cleavages by thrombin, factor Xa, and activated protein C with activation and inactivation of factor VIII coagulant activity. Biochemistry. 1986;25(2):505-512. - PubMed
-
- Vehar GA, Keyt B, Eaton D, et al. . Structure of human factor VIII. Nature. 1984;312(5992):337-342. - PubMed
-
- Lollar P, Hill-Eubanks DC, Parker CG. Association of the factor VIII light chain with von Willebrand factor. J Biol Chem. 1988;263(21):10451-10455. - PubMed
-
- Fay PJ, Smudzin TM. Characterization of the interaction between the A2 subunit and A1/A3-C1-C2 dimer in human factor VIIIa. J Biol Chem. 1992;267(19):13246-13250. - PubMed
-
- Hill-Eubanks DC, Lollar P. von Willebrand factor is a cofactor for thrombin-catalyzed cleavage of the factor VIII light chain. J Biol Chem. 1990;265(29):17854-17858. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
