

Human transfer RNA modopathies: diseases caused by aberrations in transfer RNA modifications
- PMID: 33513290
- PMCID: PMC9255597
- DOI: 10.1111/febs.15736
Human transfer RNA modopathies: diseases caused by aberrations in transfer RNA modifications
Abstract
tRNA molecules are post-transcriptionally modified by tRNA modification enzymes. Although composed of different chemistries, more than 40 types of human tRNA modifications play pivotal roles in protein synthesis by regulating tRNA structure and stability as well as decoding genetic information on mRNA. Many tRNA modifications are conserved among all three kingdoms of life, and aberrations in various human tRNA modification enzymes cause life-threatening diseases. Here, we describe the class of diseases and disorders caused by aberrations in tRNA modifications as 'tRNA modopathies'. Aberrations in over 50 tRNA modification enzymes are associated with tRNA modopathies, which most frequently manifest as dysfunctions of the brain and/or kidney, mitochondrial diseases, and cancer. However, the molecular mechanisms that link aberrant tRNA modifications to human diseases are largely unknown. In this review, we provide a comprehensive compilation of human tRNA modification functions, tRNA modification enzyme genes, and tRNA modopathies, and we summarize the elucidated pathogenic mechanisms underlying several tRNA modopathies. We will also discuss important questions that need to be addressed in order to understand the molecular pathogenesis of tRNA modopathies.
Keywords: RNA; RNA modification; tRNA; tRNA modopathy; translation.
© 2021 Federation of European Biochemical Societies.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
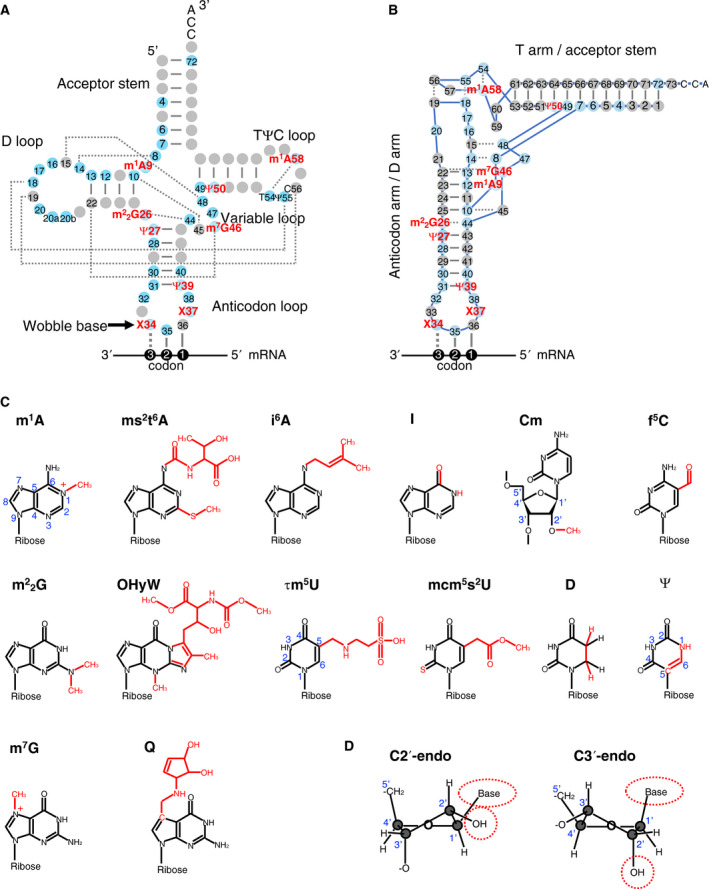
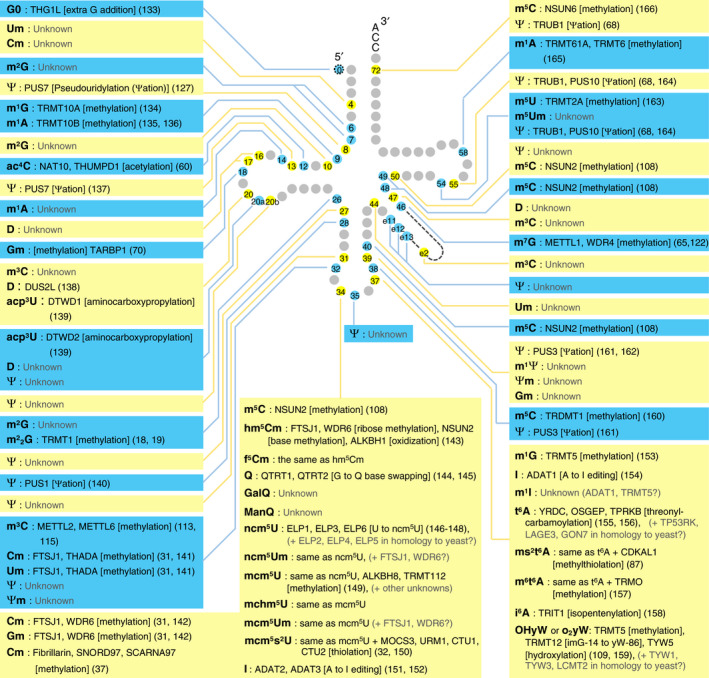
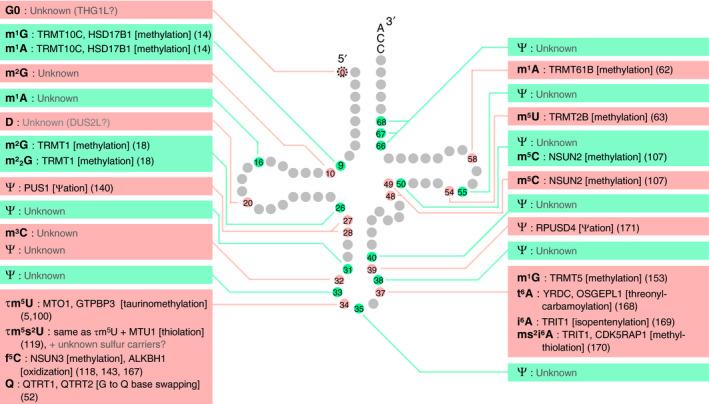
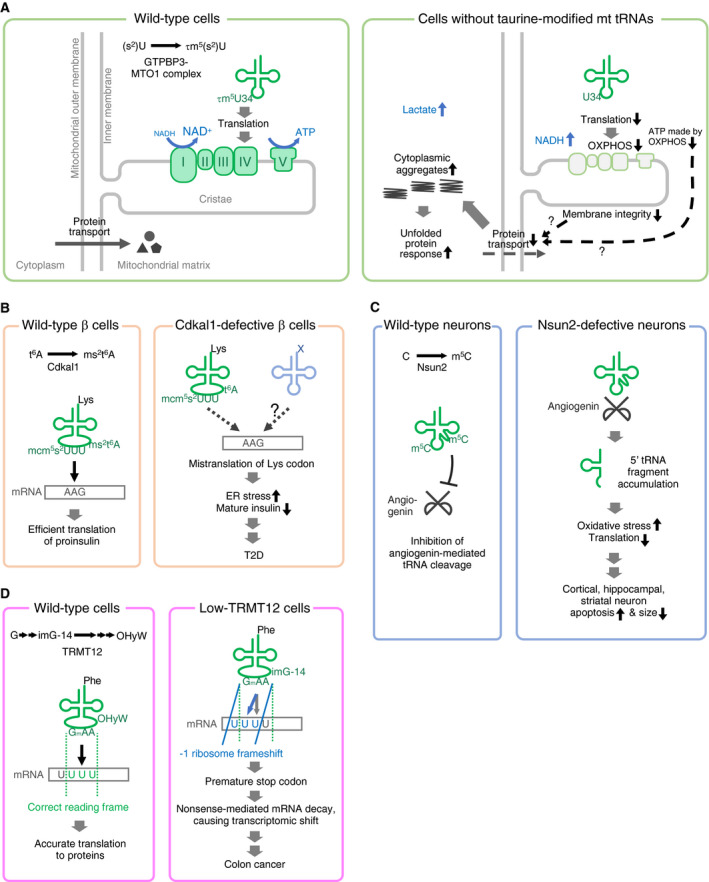
References
-
- Crick F (1958) On protein synthesis. Symp Soc Exp Biol 12, 138–163. - PubMed
-
- Hoagland MB, Stephenson ML, Scott JF, Hecht LI & Zamecnik PC (1958) A soluble ribonucleic acid intermediate in protein synthesis. J Biol Chem 231, 241–257. - PubMed
-
- Anderson S, Bankier AT, Barrell BG, de Bruijn MH, Coulson AR, Drouin J, Eperon IC, Nierlich DP, Roe BA, Sanger F et al. (1981) Sequence and organization of the human mitochondrial genome. Nature 290, 457–465. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
