

Cell-to-Cell Adhesion and Neurogenesis in Human Cortical Development: A Study Comparing 2D Monolayers with 3D Organoid Cultures
- PMID: 33513360
- PMCID: PMC7878838
- DOI: 10.1016/j.stemcr.2020.12.019
Cell-to-Cell Adhesion and Neurogenesis in Human Cortical Development: A Study Comparing 2D Monolayers with 3D Organoid Cultures
Abstract
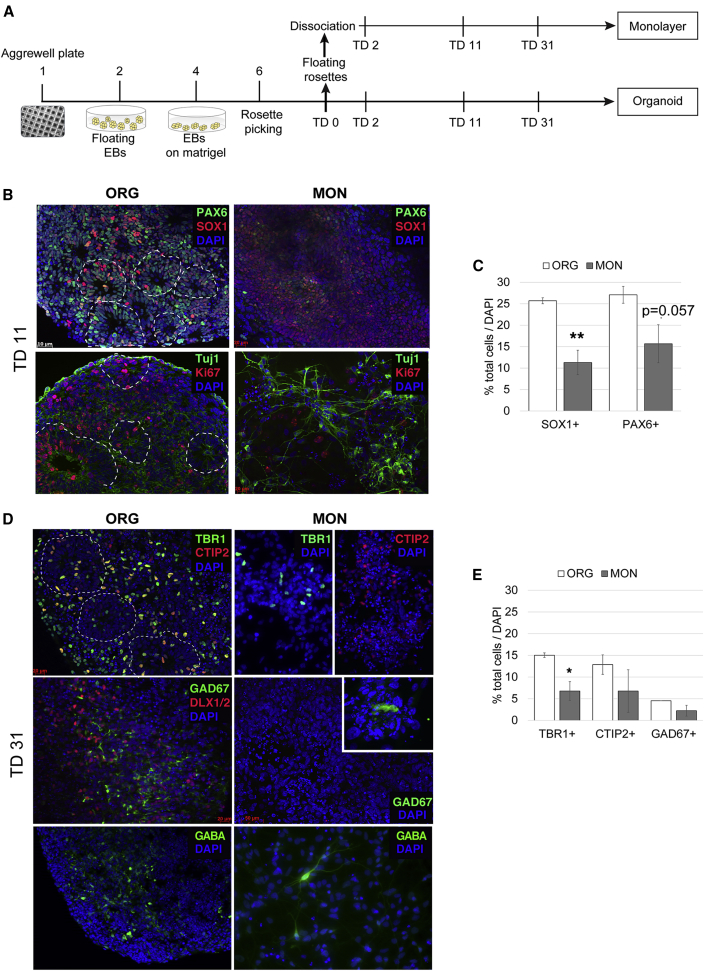
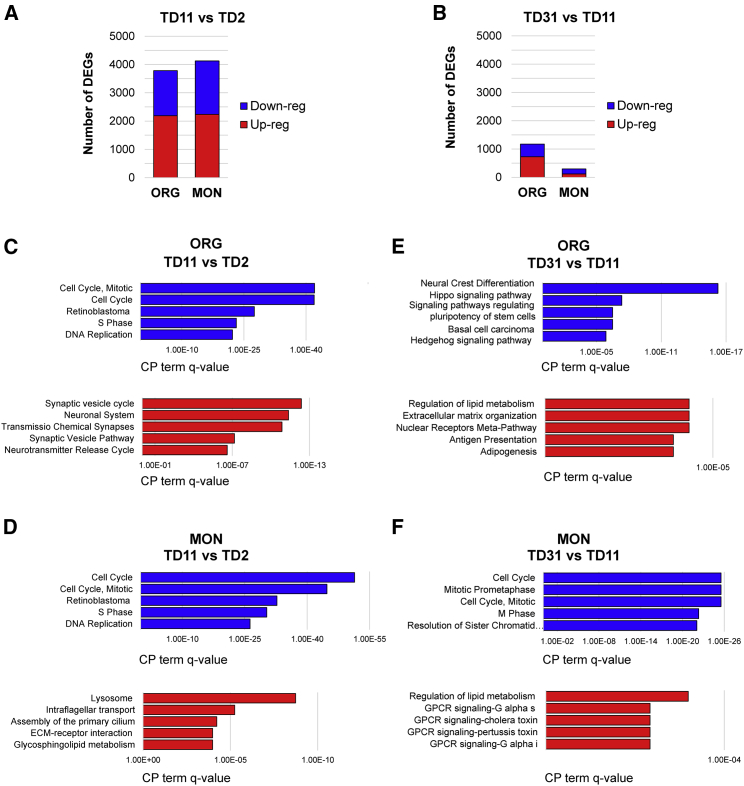
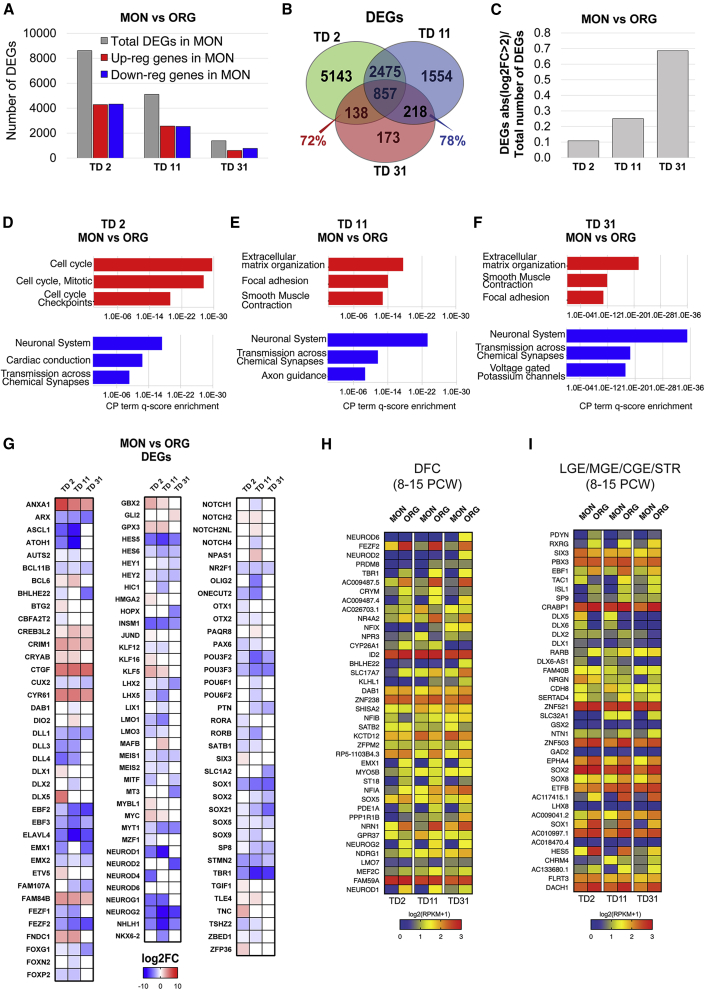
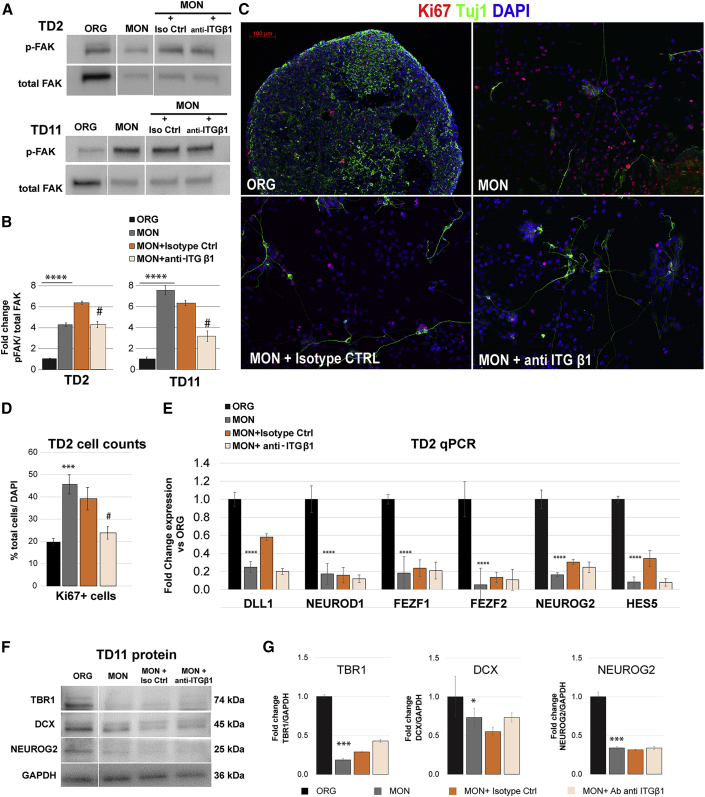
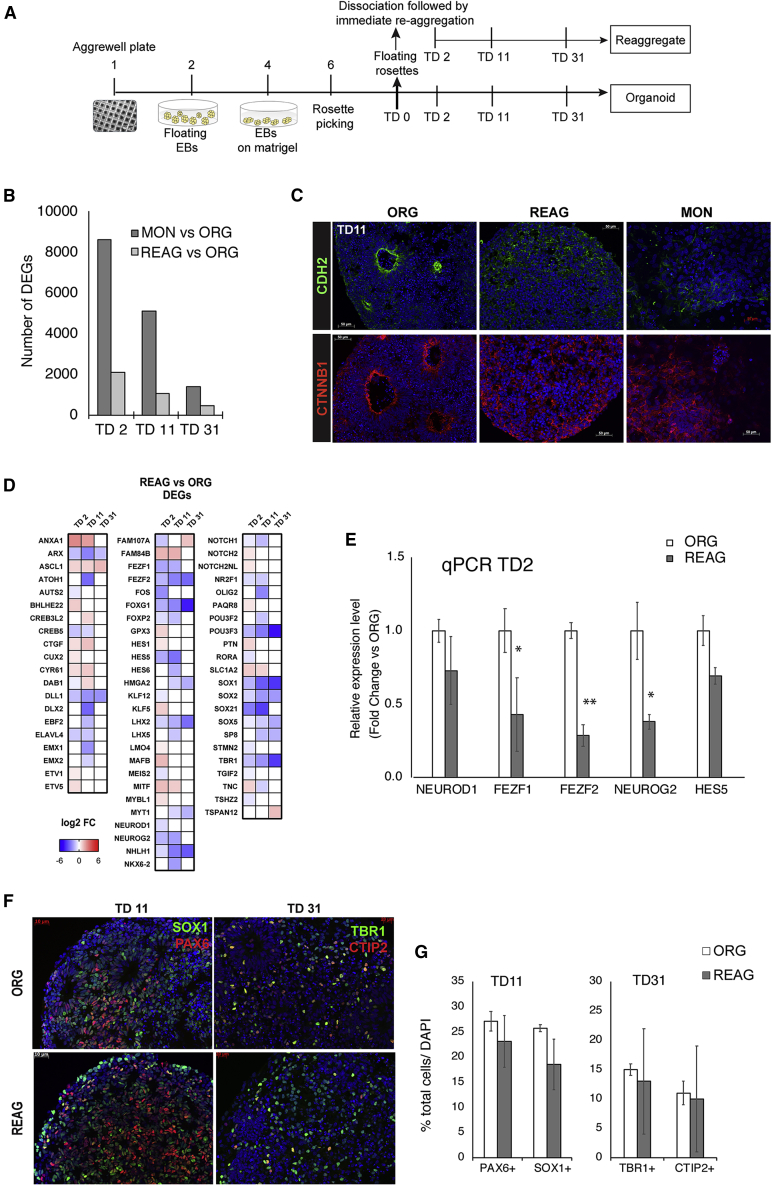
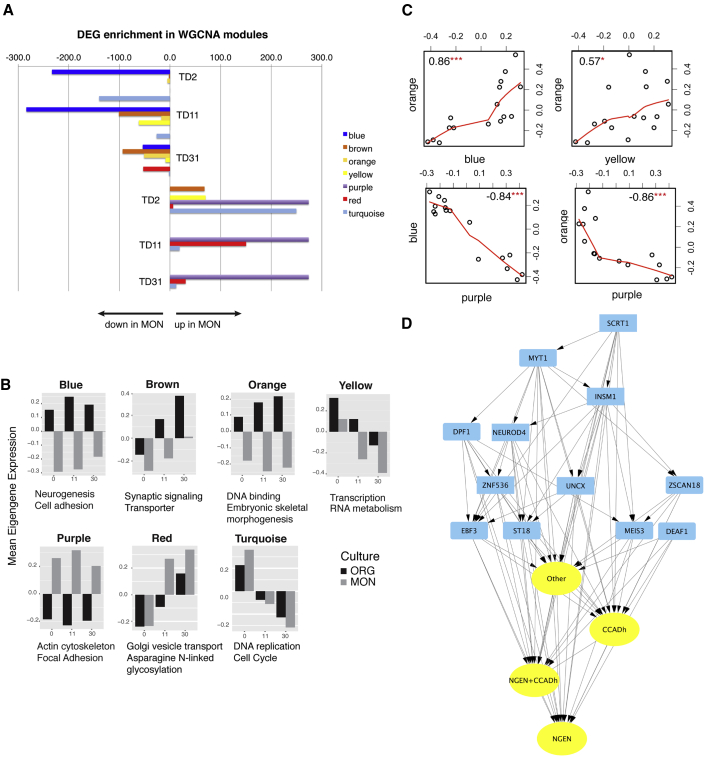
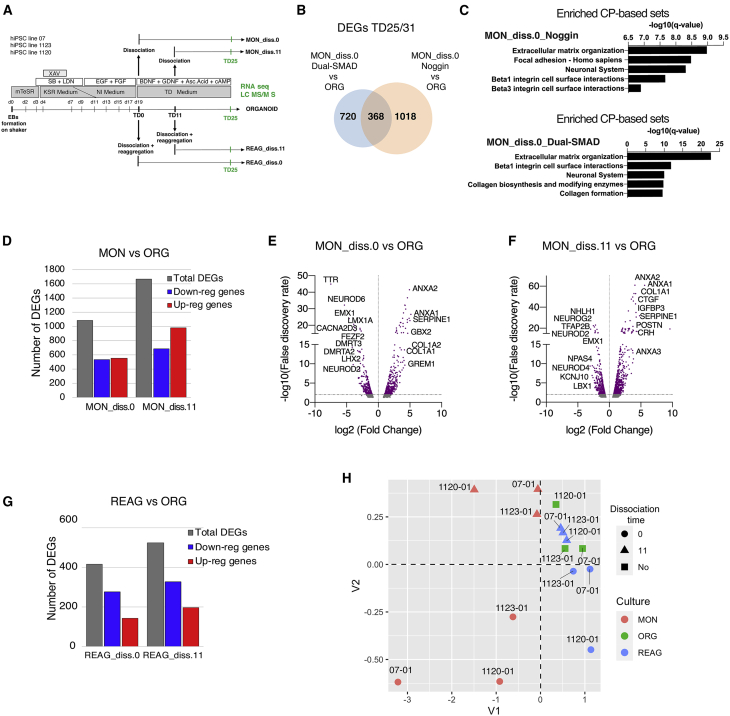
Organoids (ORGs) are increasingly used as models of cerebral cortical development. Here, we compared transcriptome and cellular phenotypes between telencephalic ORGs and monolayers (MONs) generated in parallel from three biologically distinct induced pluripotent stem cell (iPSC) lines. Multiple readouts revealed increased proliferation in MONs, which was caused by increased integrin signaling. MONs also exhibited altered radial glia (RG) polarity and suppression of Notch signaling, as well as impaired generation of intermediate progenitors, outer RG, and cortical neurons, which were all partially reversed by reaggregation of dissociated cells. Network analyses revealed co-clustering of cell adhesion, Notch-related transcripts and their transcriptional regulators in a module strongly downregulated in MONs. The data suggest that ORGs, with respect to MONs, initiate more efficient Notch signaling in ventricular RG owing to preserved cell adhesion, resulting in subsequent generation of intermediate progenitors and outer RG, in a sequence that recapitulates the cortical ontogenetic process.
Keywords: Notch signaling; RNA-seq; cerebral cortex; human; iPSCs; network analyses; organoids; proteomics.
Copyright © 2021 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Arlotta P., Pasca S.P. Cell diversity in the human cerebral cortex: from the embryo to brain organoids. Curr. Opin. Neurobiol. 2019;56:194–198. - PubMed
-
- Cappello S., Attardo A., Wu X., Iwasato T., Itohara S., Wilsch-Brauninger M., Eilken H.M., Rieger M.A., Schroeder T.T., Huttner W.B. The Rho-GTPase cdc42 regulates neural progenitor fate at the apical surface. Nat. Neurosci. 2006;9:1099–1107. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
