

Stronger induction of trained immunity by mucosal BCG or MTBVAC vaccination compared to standard intradermal vaccination
- PMID: 33521699
- PMCID: PMC7817864
- DOI: 10.1016/j.xcrm.2020.100185
Stronger induction of trained immunity by mucosal BCG or MTBVAC vaccination compared to standard intradermal vaccination
Abstract
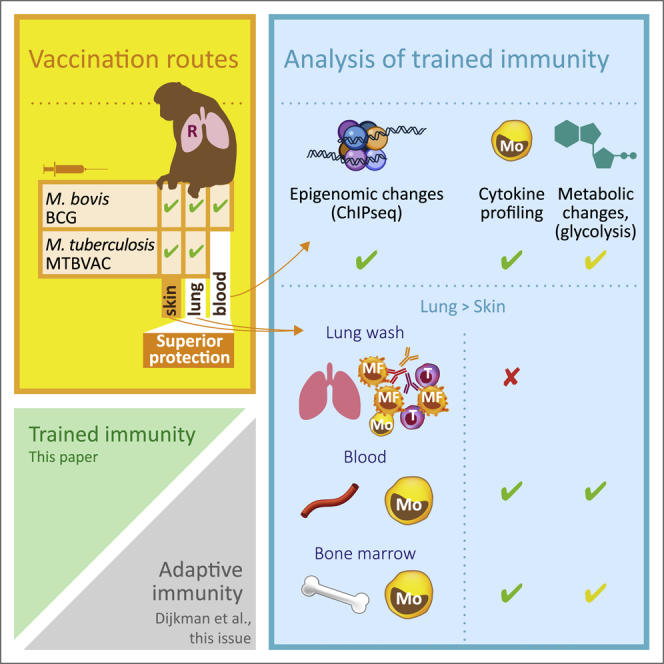
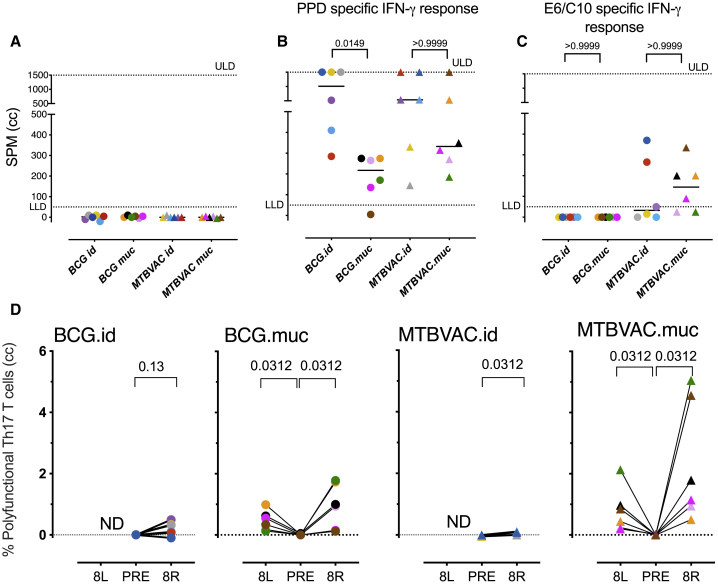
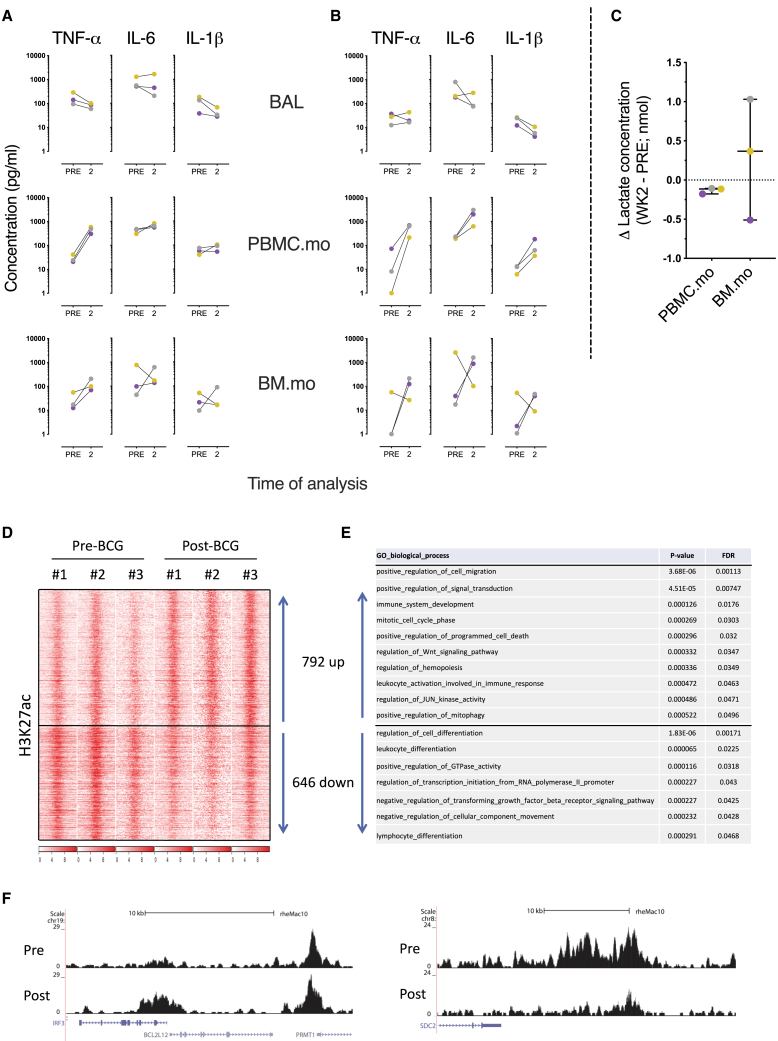
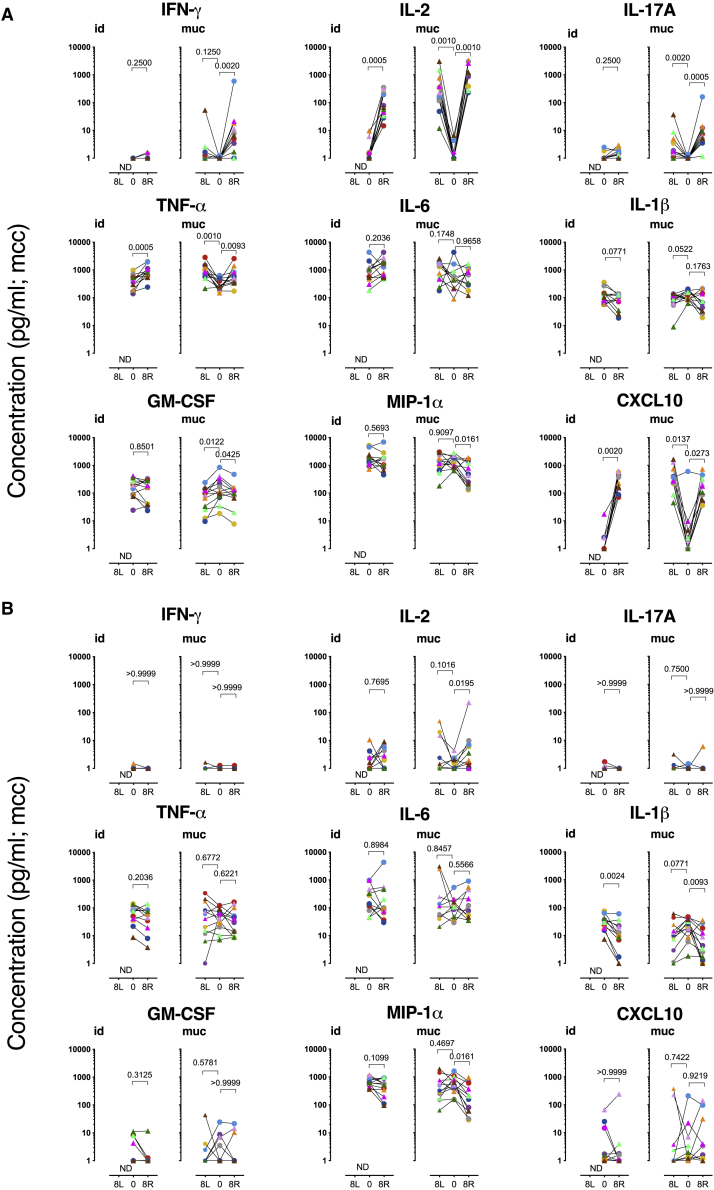
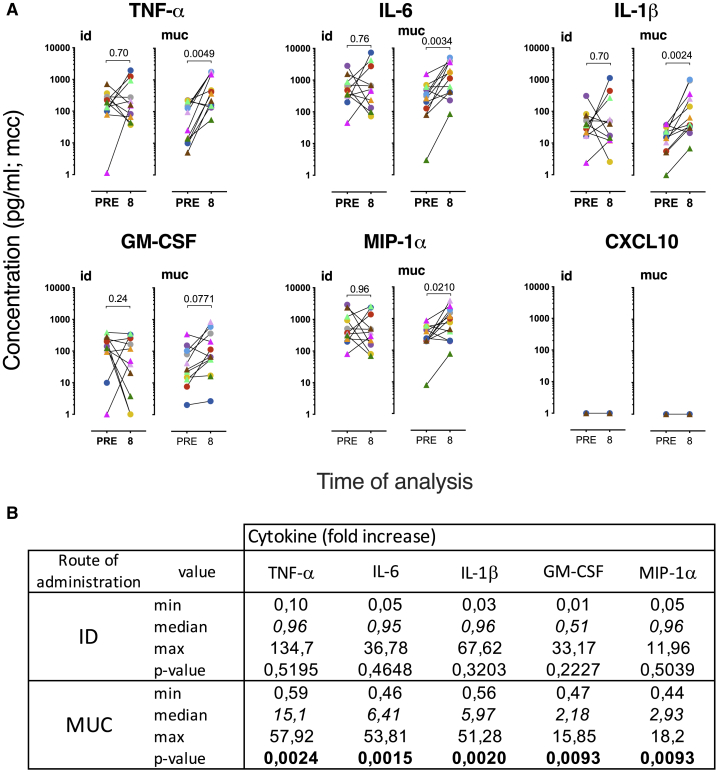
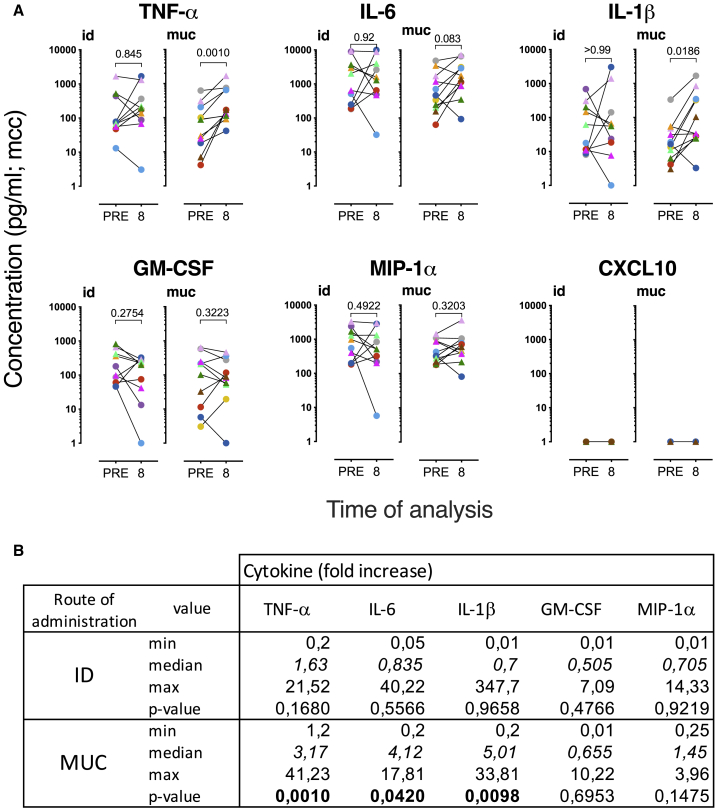
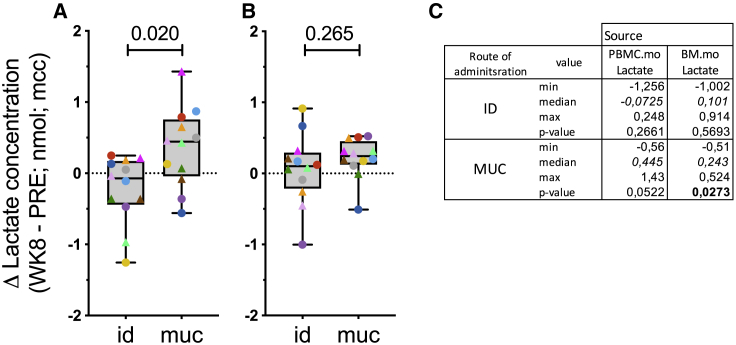
BCG vaccination can strengthen protection against pathogens through the induction of epigenetic and metabolic reprogramming of innate immune cells, a process called trained immunity. We and others recently demonstrated that mucosal or intravenous BCG better protects rhesus macaques from Mycobacterium tuberculosis infection and TB disease than standard intradermal vaccination, correlating with local adaptive immune signatures. In line with prior mouse data, here, we show in rhesus macaques that intravenous BCG enhances innate cytokine production associated with changes in H3K27 acetylation typical of trained immunity. Alternative delivery of BCG does not alter the cytokine production of unfractionated bronchial lavage cells. However, mucosal but not intradermal vaccination, either with BCG or the M. tuberculosis-derived candidate MTBVAC, enhances innate cytokine production by blood- and bone marrow-derived monocytes associated with metabolic rewiring, typical of trained immunity. These results provide support to strategies for improving TB vaccination and, more broadly, modulating innate immunity via mucosal surfaces.
Keywords: BCG; MTBVAC; immunotherapy; innate immunity; non-human primates; trained immunity; tuberculosis; vaccination; vaccine development.
© 2020 The Author(s).
Conflict of interest statement
E.R., E.P., N.A., and C.M. are co-inventors on a patent on a tuberculosis vaccine held by the University of Zaragoza and Biofabri. E.R. and E.P. are employees of Biofabri. J.T. is an employee of the TuBerculosis Vaccine Initiative and an advisor to Biofabri. M.G.N. holds a patent on the inhibition of trained immunity with nanobiologics, and a patent on the stimulation of trained immunity with nanobiologics. M.G.N. is also a scientific founder of Trained Therapeutix and Discovery (TTxD). The remaining authors of the Radboud UMC and the authors from the BPRC declare no competing interests.
Figures

References
-
- Kleinnijenhuis J., Quintin J., Preijers F., Joosten L.A., Ifrim D.C., Saeed S., Jacobs C., van Loenhout J., de Jong D., Stunnenberg H.G. Bacille Calmette-Guerin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proc. Natl. Acad. Sci. USA. 2012;109:17537–17542. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
