

Development of optically controlled "living electrodes" with long-projecting axon tracts for a synaptic brain-machine interface
- PMID: 33523957
- PMCID: PMC10670819
- DOI: 10.1126/sciadv.aay5347
Development of optically controlled "living electrodes" with long-projecting axon tracts for a synaptic brain-machine interface
Abstract
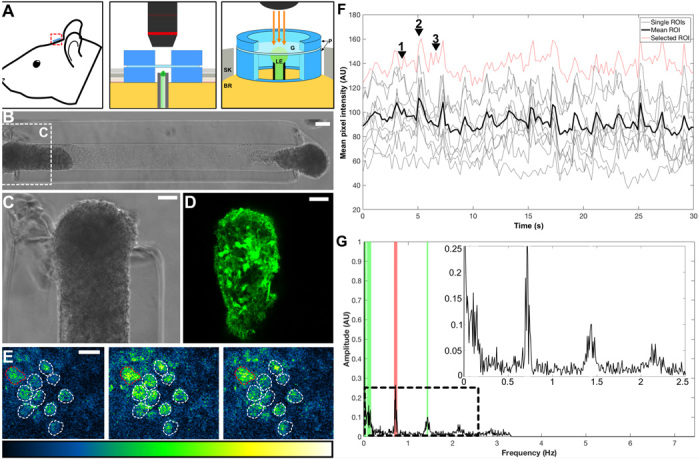
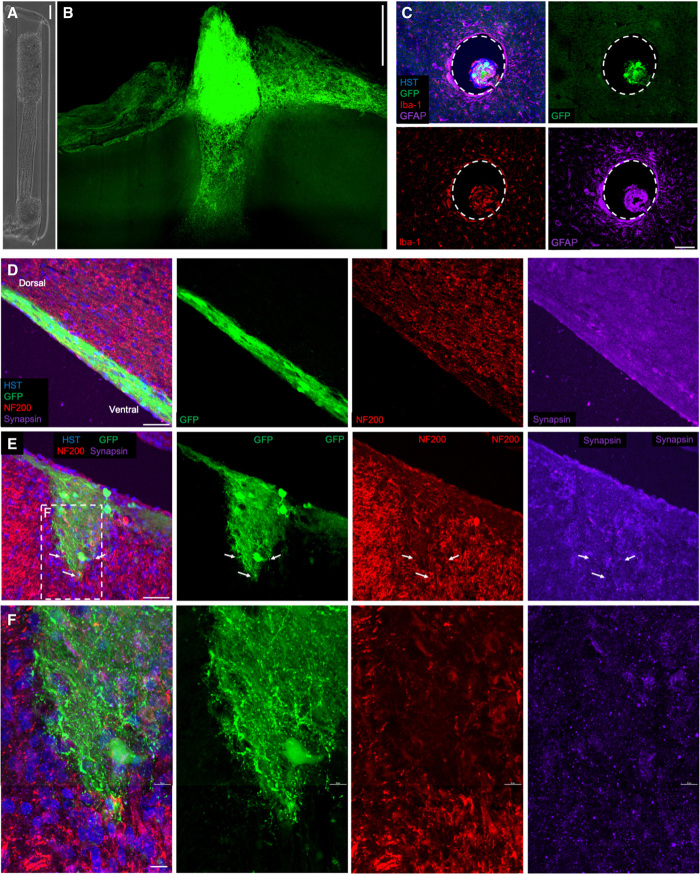
For implantable neural interfaces, functional/clinical outcomes are challenged by limitations in specificity and stability of inorganic microelectrodes. A biological intermediary between microelectrical devices and the brain may improve specificity and longevity through (i) natural synaptic integration with deep neural circuitry, (ii) accessibility on the brain surface, and (iii) optogenetic manipulation for targeted, light-based readout/control. Accordingly, we have developed implantable "living electrodes," living cortical neurons, and axonal tracts protected within soft hydrogel cylinders, for optobiological monitoring/modulation of brain activity. Here, we demonstrate fabrication, rapid axonal outgrowth, reproducible cytoarchitecture, and simultaneous optical stimulation and recording of these tissue engineered constructs in vitro. We also present their transplantation, survival, integration, and optical recording in rat cortex as an in vivo proof of concept for this neural interface paradigm. The creation and characterization of these functional, optically controllable living electrodes are critical steps in developing a new class of optobiological tools for neural interfacing.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution License 4.0 (CC BY).
Figures
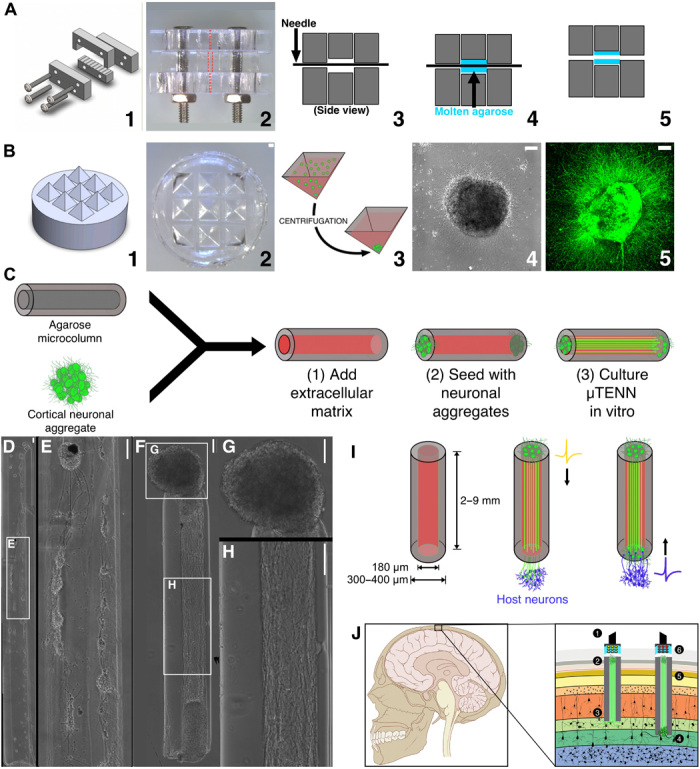
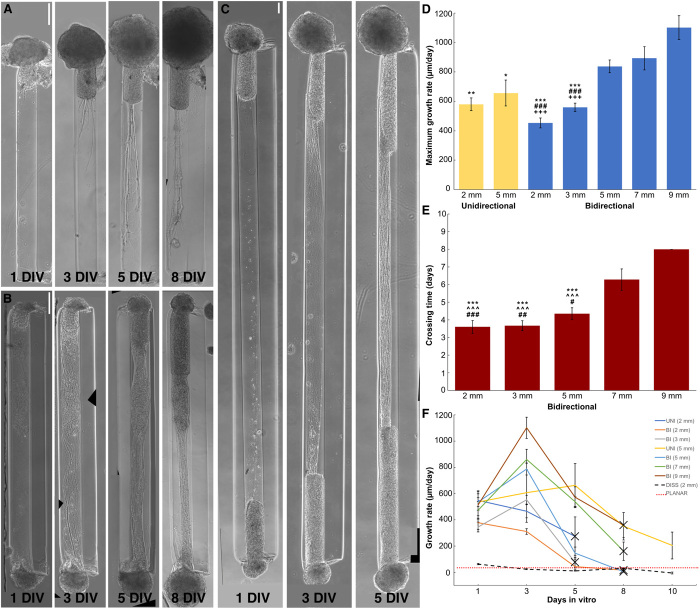
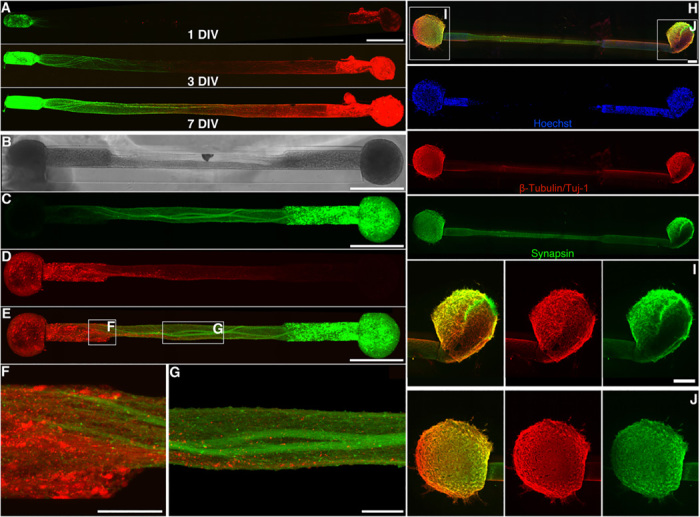
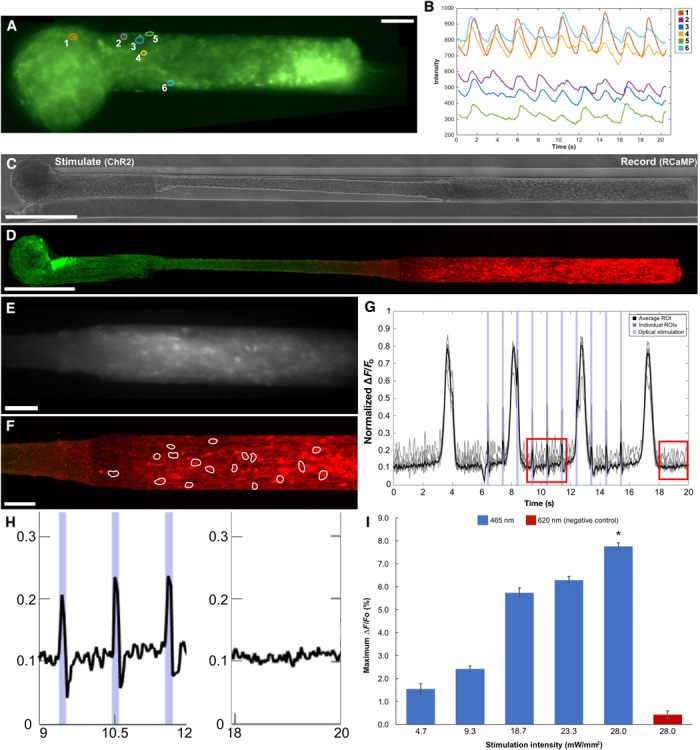
References
-
- Tresco P. A., Winslow B. D., The challenge of integrating devices into the central nervous system. Crit. Rev. Biomed. Eng. 39, 29–44 (2011). - PubMed
-
- Harris J. P., Tyler D. J., Biological, mechanical, and technological considerations affecting the longevity of intracortical electrode recordings. Crit. Rev. Biomed. Eng. 41, 435–456 (2013). - PubMed
-
- Grill W. M., Norman S. E., Bellamkonda R. V., Implanted neural interfaces: Biochallenges and engineered solutions. Annu. Rev. Biomed. Eng. 11, 1–24 (2009). - PubMed
-
- Polikov V. S., Tresco P. A., Reichert W. M., Response of brain tissue to chronically implanted neural electrodes. J. Neurosci. Methods 148, 1–18 (2005). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
