

Review
doi: 10.1101/cshperspect.a040600.
Signaling and Metabolism
Affiliations
- PMID: 33526508
- PMCID: PMC7849339
- DOI: 10.1101/cshperspect.a040600
Item in Clipboard
Review
Signaling and Metabolism
Cold Spring Harb Perspect Biol.
.
No abstract available
Figures
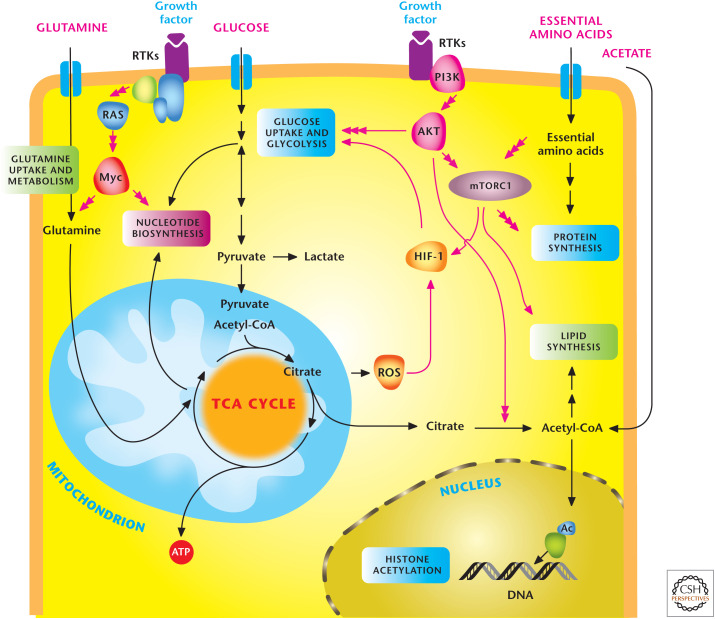
Signal transduction pathways regulate metabolism. In multicellular organisms, cells use growth factors to uptake nutrients. Growth factors through receptor tyrosine kinases (RTKs) activate the PI3K to stimulate AKT and mTORC1, which promotes glycolysis and lipid synthesis. mTORC1 signaling responds to levels of essential amino acids to stimulate protein synthesis. The HIF-1 transcription factor further promotes glycolysis. RTK activation also activates Myc transcription factor, which enhances both glycolysis and mitochondrial metabolism. Myc stimulates glutamine metabolism necessary to sustain mitochondrial metabolism and nucleotide biosynthesis. Mitochondrial ROS and acetyl-CoA production promote HIF-1 activation and histone acetylation, respectively, as examples of metabolism feeding back on signaling. (Adapted, with permission, from Ward and Thompson 2012.)
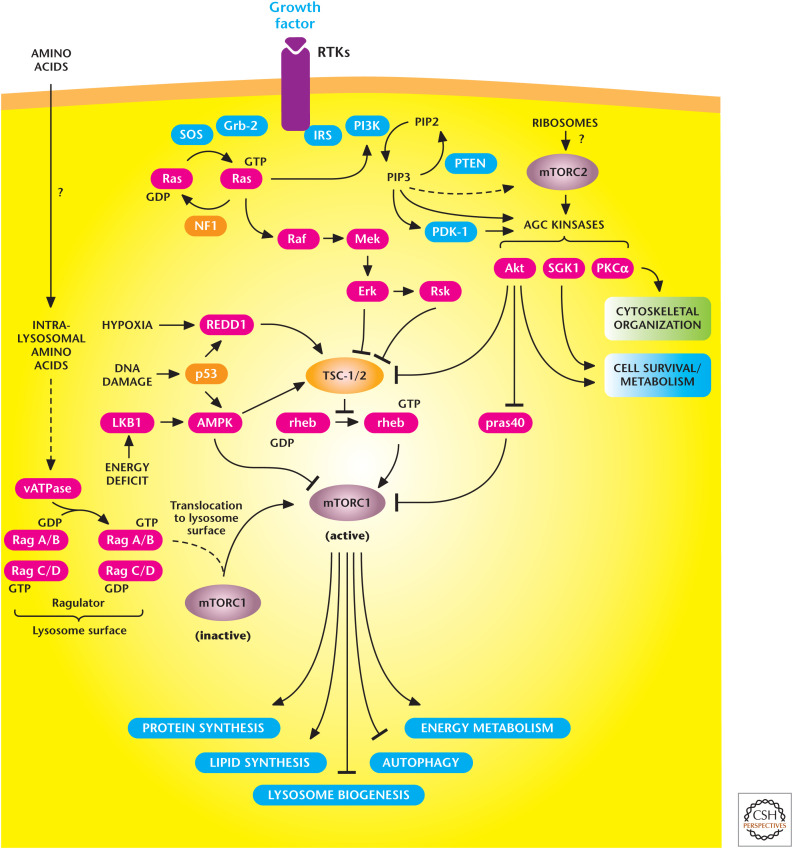
The mechanistic target of rapamycin (mTOR) signaling pathway controls metabolism and cell survival. mTORC1 stimulates multiple anabolic metabolic pathways (lipid and protein synthesis) in the presence of nutrients and growth factors. Conversely, limiting nutrient or growth factor availability promotes catabolic pathways (autophagy). mTORC2 promotes cell survival through activation of AKT. (Adapted from Laplante and Sabatini 2012.)
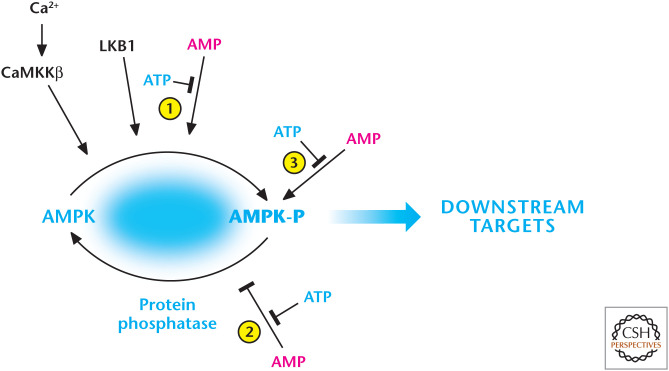
AMP and calcium activate AMP-activated protein kinase (AMPK). AMP displaces ATP on the AMPK-γ subunit to (1) promote phosphorylation and (2) inhibit dephosphorylation of threonine-172, resulting in robust activation of AMPK. The constitutively active kinase LKB1 is required for phosphorylation of threonine-172. AMP further activates the phosphorylated AMPK (3). AMPK can also be activated by threonine-172 phosphorylation catalyzed by CaMKKβ, which also activates AMPK at threonine-172 through an increase in intracellular calcium, but independent of AMP and LKB1. (Adapted from Hardie and Alessi 2013.)
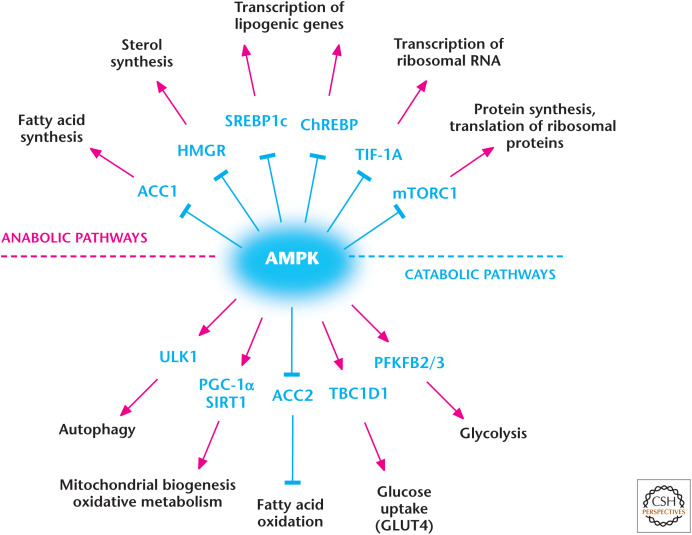
AMP-activated protein kinase (AMPK) regulates metabolism. AMPK positively and negatively regulates catabolic and anabolic pathways, respectively. (Adapted from Hardie and Alessi 2013.)

Overview of autophagy. The autophagy pathway has multiple steps, including an autophagosome that contains damaged organelles that are degraded in the final stage. This ensures organelle quality control and can generate metabolites for ATP production during nutrient-limiting conditions. (Adapted, with permission, from Choi et al. 2013, © Massachusetts Medical Society.)
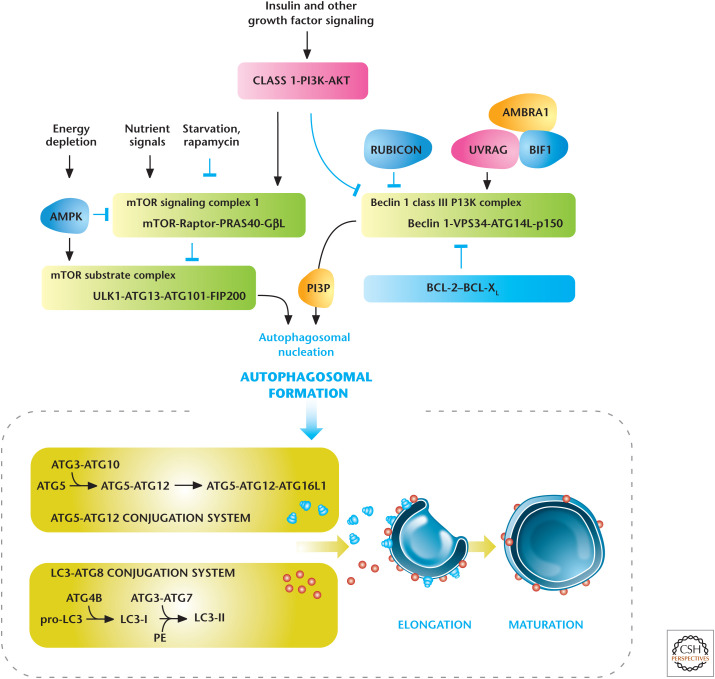
Regulators of autophagy. Growth factors and nutrients through PI3K/AKT and mTORC1, respectively, negatively regulate autophagy. AMP-activated protein kinase (AMPK) positively regulates autophagy. Autophagy is also regulated by the Beclin 1 interacting complex. Autophagosomal elongation requires the ATG5-ATG12 and LC3-ATG8 ubiquitin-like conjugation systems. In mammals, the conversion of LC3-I (free-form) to LC3-II (phosphoethanolamine-conjugated form) is used as a marker of autophagy. (Adapted, with permission, from Choi et al. 2013, © Massachusetts Medical Society.)
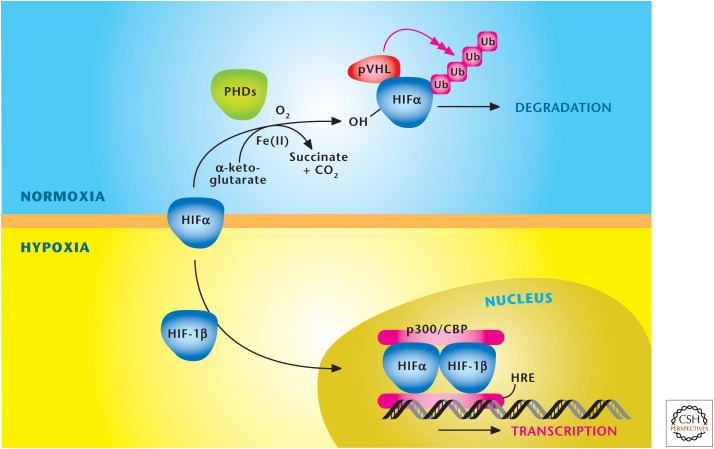
Oxygen levels control stabilization of HIFs. HIFs are a heterodimer, consisting of a constitutively stable HIF-1β and an oxygen-sensitive HIFα subunit. The HIFα subunit is hydroxylated at proline residues in the ODD by PHDs under normoxia. In addition to oxygen, the PHDs require iron (Fe2+) and α-ketoglutrate as substrates for hydroxylation. The von Hippel–Lindau protein (pVHL) recognizes hydroxylated proline residues and targets the HIFα subunit for rapid degradation by the proteasome. Hypoxia diminishes hydroxylation of HIFα to prevent its degradation and promote dimerization with HIF-1β to bind specific hypoxia response elements (HRE) in the promoter regions of its target genes. (Adapted, with permission, from Balligand et al. 2009, © The American Physiological Society.)

HIFs regulate adaptation to hypoxia. Multiple inputs, including hypoxia, ROS, TCA metabolites (succinate and fumarate), and NO, inhibit prolyl hydroxlases (PHDs) to activate hypoxia inducible factors (HIFs). At the organismal level, HIFs stimulate angiogenesis, erythropoiesis, and ventilation to increase oxygen delivery. At the cellular level, HIFs diminish oxidative phosphorylation and increase glycolysis. HIFs also suppress ATP-consuming generation to preserve a high ATP/ADP ratio in cells. (Adapted from Kaelin and Ratcliffe 2008.)
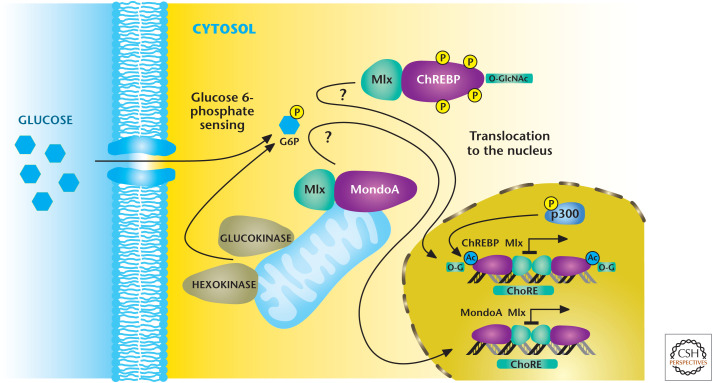
Mondo transcription factors respond to glucose flux. Mondo transcription factors ChREBP and MondoA, with their common binding partner Mlx, are stimulated through unknown mechanisms to an increase in glucose 6-phosphate levels. Subsequently, MondoA/Mlx and ChREBP/Mlx complexes translocate to the nucleus where they bind to ChoRE, resulting in activation of genes that encode enzymes in glycolysis and lipid synthesis. (Adapted from Havula and Hietakangas 2012, with permission from Elsevier.)
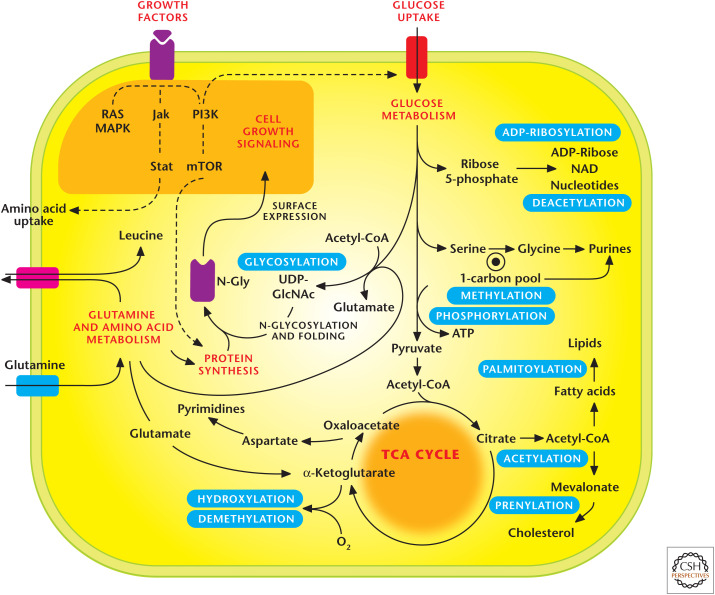
Metabolic pathways regulate signaling pathways. Metabolic pathways generate substrates that are used as posttranslational modifications (PTMs) to control signal transduction. Notable PTMs highlighted include glycosylation, prenylation, palmitoylation, hydroxylation, methylation/demethylation, phoshorylation, acetylation/deacyetylation, and ADP-ribosylation. (Adapted, with permission, from Metallo and Vander Heiden 2010.)
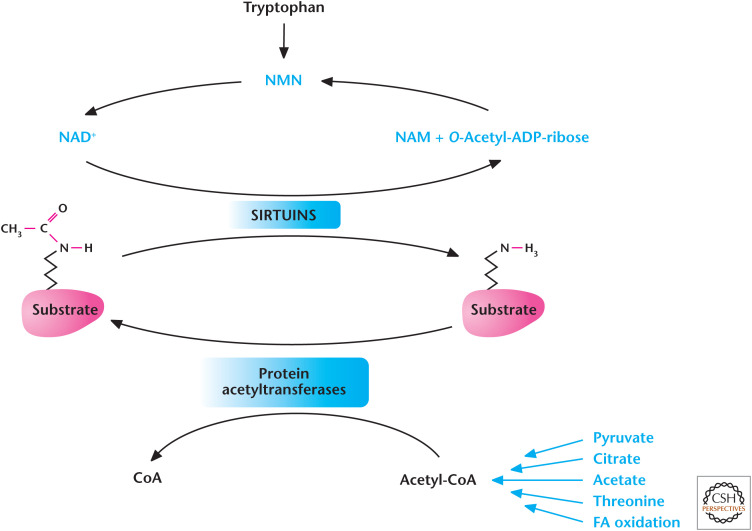
Metabolism regulates acetylation and deacetylation. Acetyl-CoA provides the acetyl group for acetylation reactions catalyzed by protein acetyltransferases. The Sirtuin family of deacetylases uses NAD+ to remove acetyl group from proteins. (Adapted from Kaelin and McKnight 2013.)
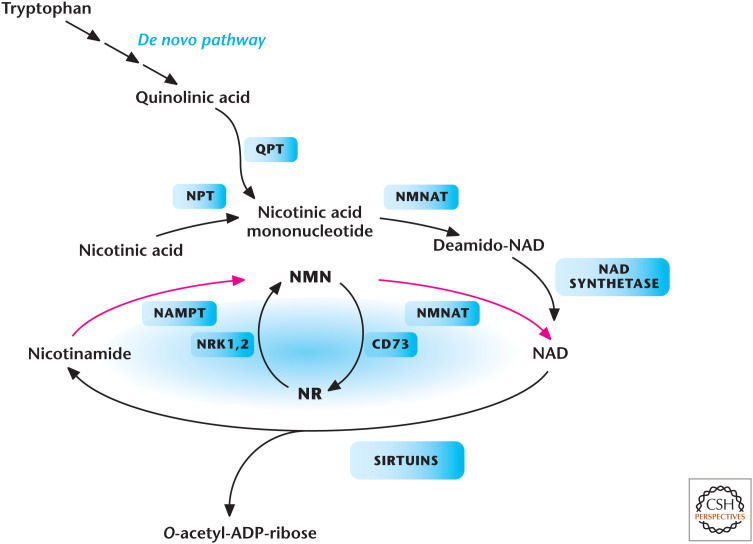
Metabolic pathways that generate NAD+ in mammals. Tryptophan, nicotinic acid, nicotinamide (NAM), or nicotinamide riboside (NR) are all NAD+ precursors. Enzymes in the pathways are quinolinate phosphoribosyltransferase (QPT), NMNAT, nicotinamide riboside kinase 1 and (NRK1,2), NAMPT, nicotinic acid phosphoribosyltransferase (NPT), and ecto-5′-nucleotidase (CD73). (Adapted from Imai and Guarente 2014, with permission from Elsevier.)

Metabolism regulates methylation. Methionine donates a methyl group to generate S-adenosylmethionine (SAM). Methyltransferases use SAM, resulting in the generation of S-adenosylhomocysteine (SAH), which is converted to homocysteine. Carbons derived from either choline- or folate-dependent reactions convert homocysteine back to methionine. DHF, dihydrofolate; THF, tetrahydrofolate; 5,10-MTHF, 5,10-methylene THF; CH3, methyl. (Adapted from Kaelin and McKnight 2013.)
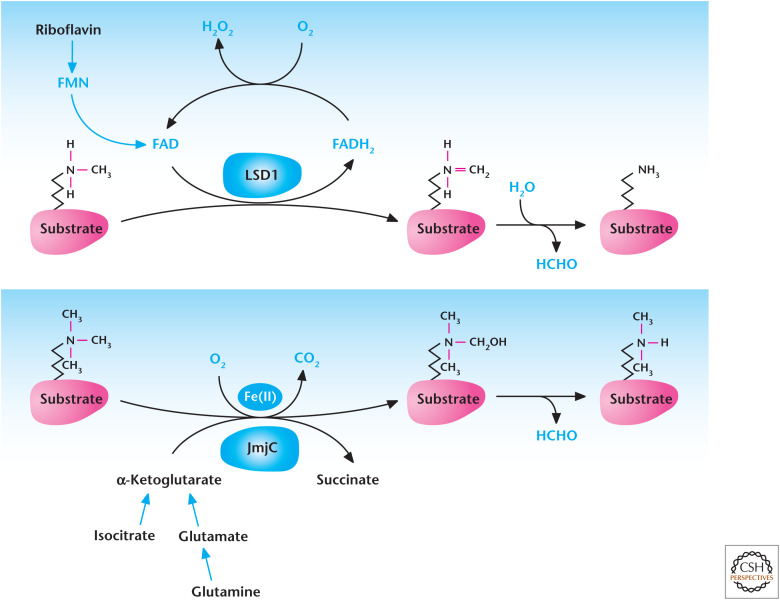
Metabolism regulates demethylation. Lysine-specific demethylases (LSD) couple oxidation of methyl groups in histones to reduction of FAD to FADH2, which spontaneously causes demethylation of histone and produces formaldehyde (HCHO). JmjC demethylases use α-ketoglutarate, Fe(II), and oxygen to hydroxylate methylated histones resulting in their demethylation.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources