

Histone H3Q5 serotonylation stabilizes H3K4 methylation and potentiates its readout
- PMID: 33526675
- PMCID: PMC8017887
- DOI: 10.1073/pnas.2016742118
Histone H3Q5 serotonylation stabilizes H3K4 methylation and potentiates its readout
Abstract
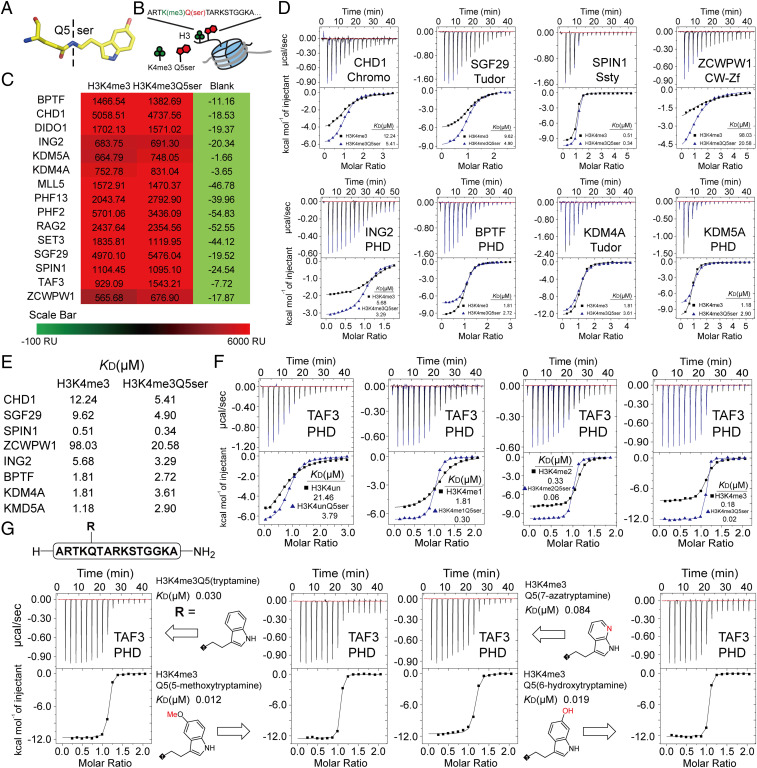
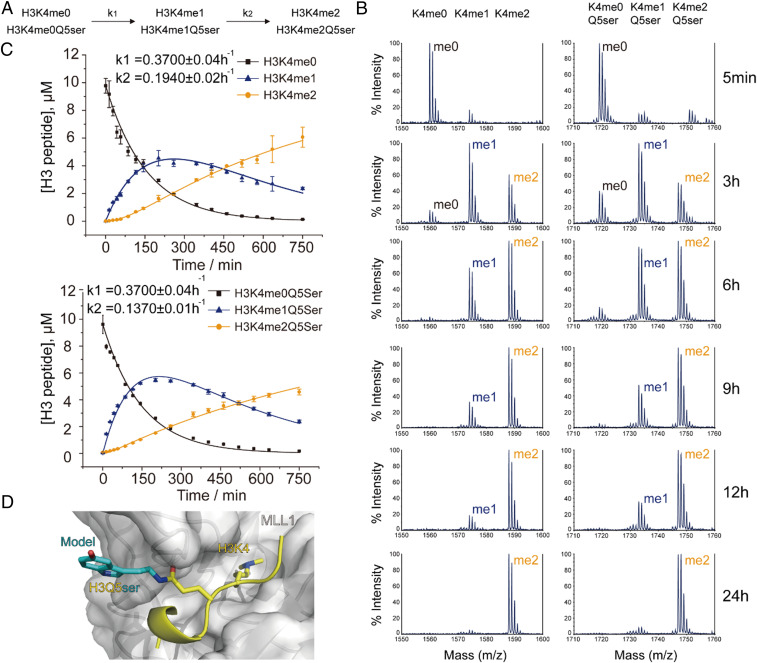
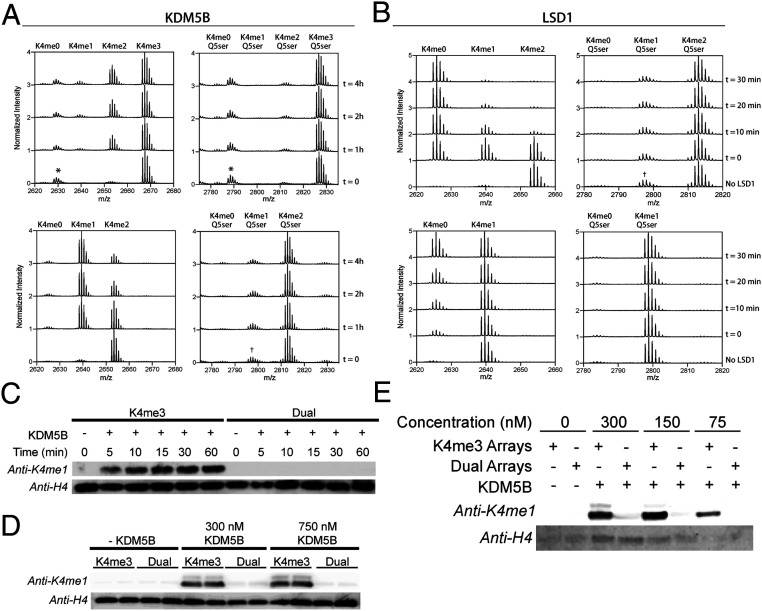
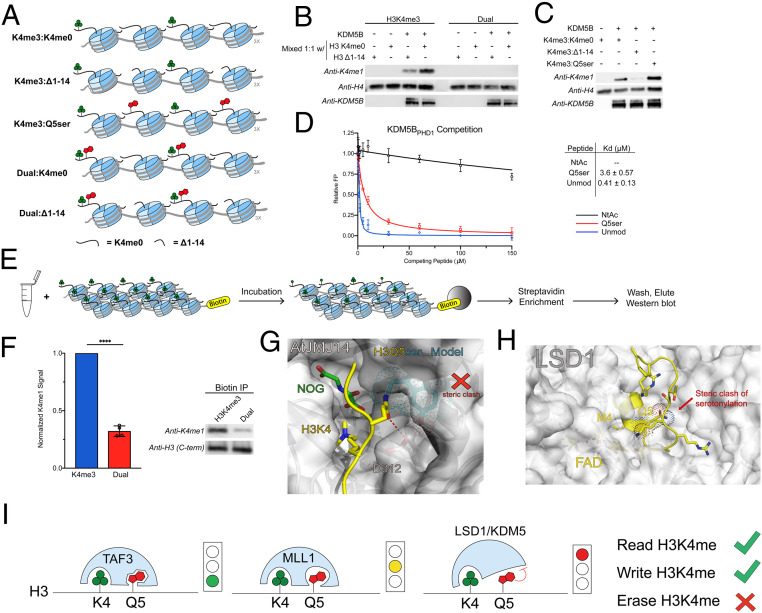
Serotonylation of glutamine 5 on histone H3 (H3Q5ser) was recently identified as a permissive posttranslational modification that coexists with adjacent lysine 4 trimethylation (H3K4me3). While the resulting dual modification, H3K4me3Q5ser, is enriched at regions of active gene expression in serotonergic neurons, the molecular outcome underlying H3K4me3-H3Q5ser crosstalk remains largely unexplored. Herein, we examine the impact of H3Q5ser on the readers, writers, and erasers of H3K4me3. All tested H3K4me3 readers retain binding to the H3K4me3Q5ser dual modification. Of note, the PHD finger of TAF3 favors H3K4me3Q5ser, and this binding preference is dependent on the Q5ser modification regardless of H3K4 methylation states. While the activity of the H3K4 methyltransferase, MLL1, is unaffected by H3Q5ser, the corresponding H3K4me3/2 erasers, KDM5B/C and LSD1, are profoundly inhibited by the presence of the mark. Collectively, this work suggests that adjacent H3Q5ser potentiates H3K4me3 function by either stabilizing H3K4me3 from dynamic turnover or enhancing its physical readout by downstream effectors, thereby potentially providing a mechanism for fine-tuning critical gene expression programs.
Keywords: H3K4me3; H3Q5 serotonylation; designer chromatin; histone modification; modification crosstalk.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
