

Mitochondrial translation deficiency impairs NAD+ -mediated lysosomal acidification
- PMID: 33528041
- PMCID: PMC8047443
- DOI: 10.15252/embj.2020105268
Mitochondrial translation deficiency impairs NAD+ -mediated lysosomal acidification
Abstract
Mitochondrial translation dysfunction is associated with neurodegenerative and cardiovascular diseases. Cells eliminate defective mitochondria by the lysosomal machinery via autophagy. The relationship between mitochondrial translation and lysosomal function is unknown. In this study, mitochondrial translation-deficient hearts from p32-knockout mice were found to exhibit enlarged lysosomes containing lipofuscin, suggesting impaired lysosome and autolysosome function. These mice also displayed autophagic abnormalities, such as p62 accumulation and LC3 localization around broken mitochondria. The expression of genes encoding for nicotinamide adenine dinucleotide (NAD+ ) biosynthetic enzymes-Nmnat3 and Nampt-and NAD+ levels were decreased, suggesting that NAD+ is essential for maintaining lysosomal acidification. Conversely, nicotinamide mononucleotide (NMN) administration or Nmnat3 overexpression rescued lysosomal acidification. Nmnat3 gene expression is suppressed by HIF1α, a transcription factor that is stabilized by mitochondrial translation dysfunction, suggesting that HIF1α-Nmnat3-mediated NAD+ production is important for lysosomal function. The glycolytic enzymes GAPDH and PGK1 were found associated with lysosomal vesicles, and NAD+ was required for ATP production around lysosomal vesicles. Thus, we conclude that NAD+ content affected by mitochondrial dysfunction is essential for lysosomal maintenance.
Keywords: GAPDH; NAD+; Nmnat3; lysosome; mitochondria.
© 2021 The Authors. Published under the terms of the CC BY 4.0 license.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
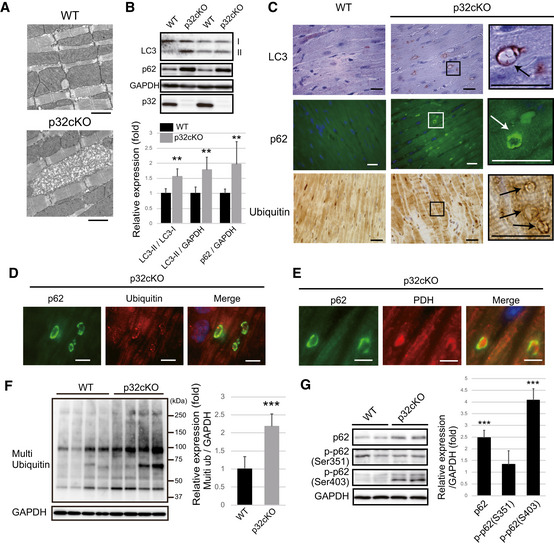
Electron microscopy of abnormal mitochondria in p32cKO hearts of 6‐month‐old mice. These two figures are also presented in Fig EV1A along with other EM figures. Scale bars, 1 µm.
Western blot analysis of autophagy marker proteins (LC3‐II and p62) in 6 months old WT and p32cKO hearts. The bands of LC3‐II and p62 are increased in the p32cKO heart. GAPDH was used as an internal control. Quantification is shown on the bottom. These values are presented as mean ± SD (n = 6 mice per group). Student’s t‐test was performed on WT vs. p32cKO, **P < 0.01.
Immunostaining of LC3, p62, and multi‐ubiquitin in heart tissues of 6 months old. The magnified image (right panel) shows the ring‐like staining pattern in the p32cKO heart (indicated arrows). Scale bar, 20 µm. The panel related to LC3 immunostaining in p32cKO is also presented in Fig EV1B (top left) along with other immunostaining pictures.
In the 6‐month‐old p32cKO heart, p62 and multi‐ubiquitin staining patterns were co‐localized. Scale bar, 5 µm.
In the 6‐month‐old p32cKO heart, the staining patterns of p62 and the mitochondrial marker protein, pyruvate dehydrogenase (PDH), were co‐localized. Scale bar, 5 µm.
Western blot analysis of ubiquitinated proteins using an anti‐multi‐ubiquitin antibody in the 6‐month‐old WT and p32cKO heart (n = 4 mice per group). Error bars are presented as mean ± SD. Student’s t‐test was performed on WT vs. p32cKO, ***P < 0.005.
Western blot analysis of phosphorylated p62 (Se351 and Ser403) and non‐phosphorylated p62 in the 6‐month‐old WT and p32cKO heart. Relative expression of phosphorylated p62 is shown on the right. Error bars are presented as mean ± SD. Student’s t‐test was performed on eight WT mice vs. seven p32cKO mice, ***P < 0.005.
- A
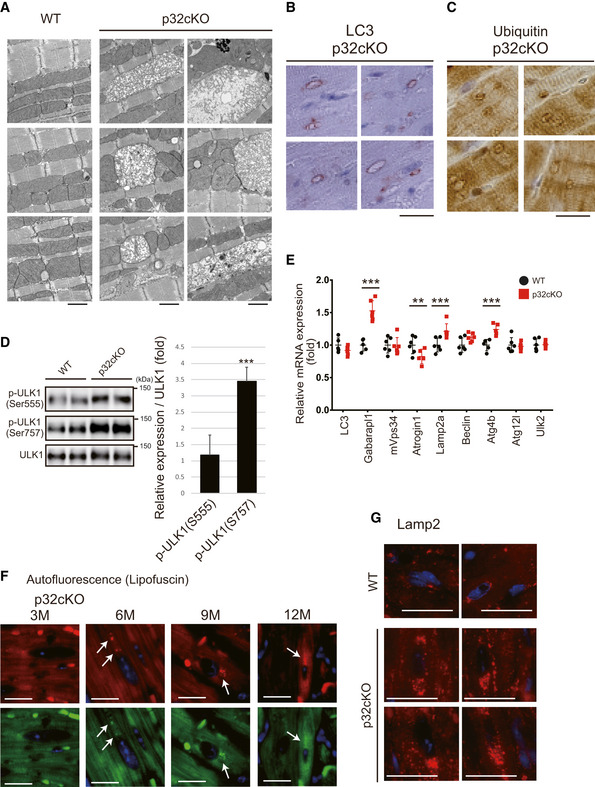
Electron microscopy of hearts from 6‐month‐old mice (n = 6). EMs of three WT and six p32cKO heart were presented. Two of them are posted as used in Fig 1A. p32cKO hearts showed morphologically abnormal mitochondria. Scale bars, 1 µm.
- B, C
Immunostaining of LC3 (B) and ubiquitin (C) in the 6‐month‐old p32cKO heart tissues. Only p32cKO hearts showed ring‐shaped structures (n = 4). Scale bar, 20 µm.
- D
Western blot analysis of phosphorylated ULK1. The p‐ULK1(Ser757) levels were higher in the 9‐month‐old p32cKO hearts than in WT hearts (n = 6 mice per group). The p‐ULK1(Ser555) levels were similar in p32cKO and WT hearts. Relative expression of phosphorylated ULK1 is shown on the right. Error bars are presented as mean ± SD. Statistical significance was assessed by Student’s t‐test, ***P < 0.005.
- E
Real‐time PCR analysis of RNA expression of autophagy‐related genes in the 6‐month‐old hearts of WT and p32cKO mice (n = 6). Error bars are presented as mean ± SD. Statistical significance was assessed by Student’s t‐test, **P < 0.01, ***P < 0.001.
- F
Autofluorescence showing lipofuscin localization around the nucleus in the p32cKO heart at different ages. Tissues were excited at a wavelength of 540 (upper panel) or 470 (lower panel) and emission spectra were collected with a confocal microscope at wavelengths (band path) of 580–630 nm (upper panel) or 510–560 nm (lower panel). Lipofuscin is indicated by arrows. Scale bars, 20 µm.
- G
Immunostaining of Lamp2 in the heart of 12‐month‐old mice using TrueBlack™ to quench lipofuscin autofluorescence. In the p32cKO heart, but not in WT, many dots around nuclei were observed. Scale bars, 20 µm.
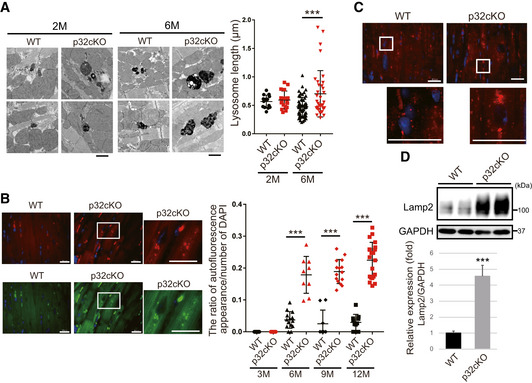
Electron micrographs showing lysosomes in cardiac myofibrils of 2 and 6 months old mice. Lysosome size and density were different in WT (black dot) and p32cKO (red dot) heart tissues. Scale bars, 1 µm. The quantification of the average lysosomal length is presented in the plot in the right panel as mean ± SEM (2‐month‐old mice, n = 14–16; 6‐month‐old mice, n = 35–60). Statistical significance was assessed by Student’s t‐test, ***P < 0.005.
Autofluorescence showing lipofuscin localization around the nucleus in the 6 months old p32cKO heart. Tissues were excited at a wavelength of 540 (upper panel) or 470 (lower panel), and emission spectra were collected with a confocal microscope at wavelengths (band path) of 580–630 nm (upper panel) or 510–560 nm (lower panel). Enlarged view of the white squares is shown right. Scale bars, 20 µm. The quantification of the ratio of autofluorescence of DAPI staining is presented in the plot in the right panel as mean ± SEM (3‐month‐old mice, n = 11; 6‐month‐old mice, n = 9–14, 9‐month‐old mice, n = 9–15, 12‐month‐old mice, n = 11–21). Statistical significance was assessed by Student’s t‐test, ***P < 0.005.
Immunostaining of Lamp2 in the heart using TrueBlack™ to quench lipofuscin autofluorescence in the 6‐month‐old WT and p32cKO heart. Enlarged view of the white squares is shown below. Scale bars, 20 µm.
Western blot analysis of Lamp2 in the 6 months old WT and p32cKO hearts. Quantification is shown in the lower panel (n = 4 mice per group). GAPDH was used as an internal control. Error bars are presented as mean ± SD. Student’s t‐test was performed on WT vs. p32cKO, ***P < 0.005.
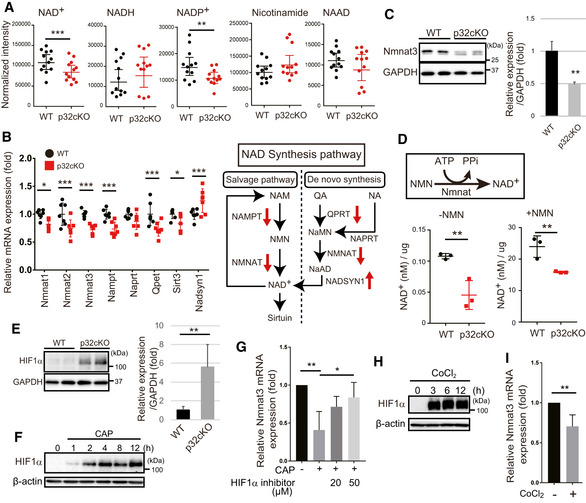
LC‐MS/MS metabolomic analysis of NAD+, NADH, NADP+, nicotinamide, and NAAD in the 6‐month‐old WT and p32cKO hearts. NAD+ and NADP+ showed significantly different levels between WT and p32cKO hearts (n = 12). Error bars are presented as mean ± SD. Student’s t‐test was performed on WT vs. p32cKO, **P < 0.001, ***P < 0.0002.
Real‐time PCR analysis of RNA expression of NAD‐synthesizing enzymes in the 6 months old WT and p32cKO hearts. Error bars are presented as mean ± SD. Student’s t‐test was performed on WT mice vs. p32cKO mice (n = 6), ***P < 0.005, *P < 0.05. The right panel shows the NAD synthesis pathway and mRNA expression levels are indicated by red arrows. NAM: nicotinamide, NA: nicotinic acid, QA: quinolic acid, NMN: nicotinamide mononucleotide.
Reduced Nmnat3 expression in 9‐month‐old p32cKO heart. Quantification is shown on the right (n = 4 mice per group). Error bars are presented as mean ± SD. Student’s t‐test was performed on WT vs. p32cKO, **P < 0.01.
NAD+ levels without or with NMN addition in 9‐month‐old WT and p32cKO heart lysates (n = 3). Error bars are presented as mean ± SD. Student’s t‐test was performed on WT vs. p32cKO, **P < 0.002.
Western blot analysis of HIF1α in 9‐month‐old WT and p32cKO heart. Quantifications is shown on the right (n = 4 mice per group). Error bars are presented as mean ± SD. Student’s t‐test was performed on WT vs. p32cKO, **P < 0.01. GAPDH was used as an internal control.
Immunoblot analysis of HIF1α; the expression in 3T3‐L1 cells was increased after treatment with 1 mM chloramphenicol (CAP), which inhibits mitochondrial translation. β‐actin was used as an internal control. One representative experiment out of three shown.
Nmnat3 mRNA expression in 3T3‐L1 cells after 1 mM CAP treatment for 72 h. The HIF1α inhibitor (20 or 50 µM) was added 2 h before the CAP treatment. Error bars are presented as mean ± SD of three independent experiments. Student’s t‐test was performed on WT cells vs. WT cells treated CAP and (or) HIF1α inhibitor, **P < 0.01, *P < 0.05.
Immunoblot analysis of HIF1α after treatment with 150 µM CoCl2 in 3T3‐L1 cells, which stabilized HIF1α. β‐actin was used as an internal control.
Nmnat3 mRNA expression in 3T3‐L1 cells after 150 µM CoCl2 treatment for 72 h (n = 3). Error bars are presented as mean ± SD of three independent experiments. Statistical significance was assessed by Student’s t‐test, **P < 0.01.
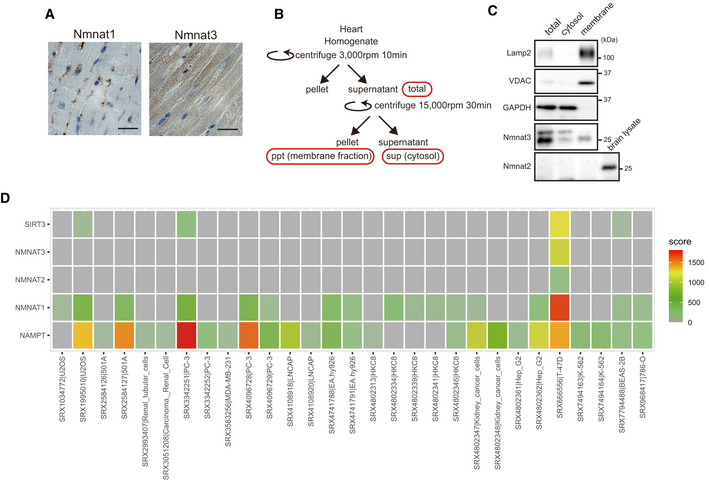
Immunostaining of Nmnat1 (left) and Nmnat3 (right) in the 9‐month‐old WT heart. Nmnat1 and Nmnat3 are localized in the nucleus and cytosol, respectively. Scale bar, 20 µm. One representative experiment out of three shown.
A schema of the procedure of membrane fraction and cytosol purification.
Western blot analysis of Nmnat3 in the 3‐month‐old WT mouse heart. Nmnat3 expression was observed in both cytosolic and membrane fractions. Lamp2 and VDAC are membrane fraction markers, and PGK1 and GAPDH are cytosolic markers. The expression of Nmnat2 was not observed in heart, but expressed in the 2‐month‐old WT mouse brain. One representative experiment out of three shown.
HIF1α was associated with the promoter region of Nampt, Nmnat1–3, and Sirt3 in several cell lines. We used the public database, ChIP‐Atlas (
http://chip‐atlas.org/ ).
Lysosomal acidification is impaired in p32KO MEFs. Representative images of WT and p32KO MEFs, stained with dextran‐Oregon Green (488) and dextran‐TMRM. Scale bar, 5 µm. Addition of 1 mM NMN for 48 h to p32KO MEFs rescued lysosomal acidification, and 1 mM CAP treatment of WT MEFs for 48 h decreased lysosomal acidification. Dextran‐Oregon Green is quenched under acidic pH, and thus, an increased green:red ratio denotes impaired lysosomal acidification. The average ± SEM of the green:red ratio in at least 30 cells in two independent experiments is presented in the plot (normalized to WT green/red = 1). Statistical significance was assessed by Student’s t‐test, *P < 0.05, ***P < 0.001.
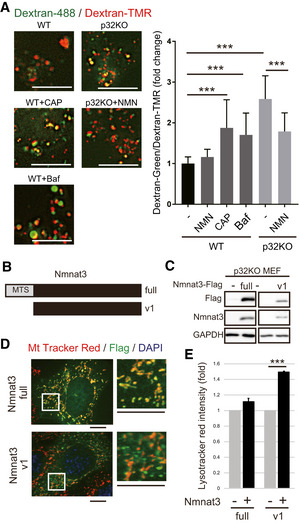
Two Nmnat3 constructs: Nmnat3 (full) with a mitochondrial‐targeting sequence (MTS), and Nmnat3(v1) without MTS.
Both these constructs were effectively expressed in p32KO MEFs. After transfection, lysates were subjected to Western blot analysis with anti‐Flag and anti‐Nmnat3 antibodies. GAPDH was used as an internal control. One representative experiment out of three shown.
Nmnat3 (full) was localized in mitochondria, but Nmnat3(v1) did not co‐stain with MitoTracker red in p32KO MEFs. Scale bars, 10 µm. The magnified image is shown on the right.
Nmnat3(v1) overexpression in p32KO MEFs rescued lysosomal function. LysoTracker Red‐stained p32KO MEFs were quantified by flow cytometry. Error bars are presented as mean ± SEM of 4 independent experiments. Statistical significance was assessed by Student’s t‐test, ***P < 0.001.
- A
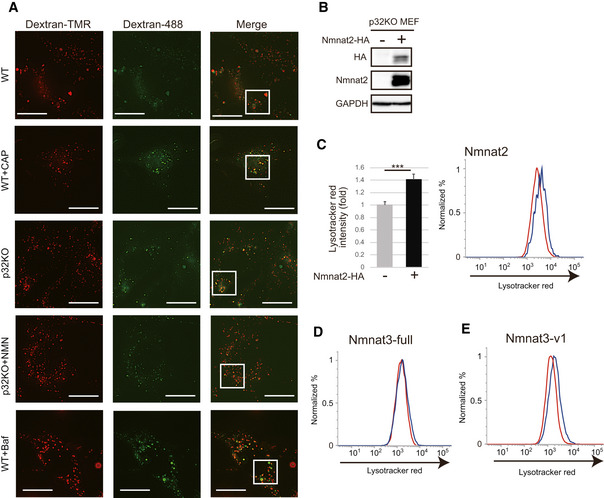
Lysosomal acidification was impaired by treatment with CAP and in p32KO MEFs. Representative images of WT and p32KO MEFs stained with dextran‐Oregon Green (488) and dextran‐TMRM. Scale bar, 5 µm. Addition of 1 mM NMN for 48 h to p32KO MEFs rescued lysosomal acidification, while 1 mM CAP treatment of WT MEFs for 48 h decreased lysosomal acidification. Enlarged view of the white squares is shown Fig 4A. One representative experiment out of three shown.
- B
Nmnat2‐HA was transfected into p32KO MEFs. Western blot analysis of anti‐HA and anti‐Nmnat2 is shown. GAPDH was used as a loading control.
- C
Overexpression of Nmnat2‐HA cells into p32KO MEFs increased LysoTracker Red staining. The light side was analyzed by FACS with LysoTracker Red. Error bars are presented as mean ± SEM of three independent experiments. Statistical significance was assessed by Student’s t‐test, ***P < 0.001.
- D, E
FACS analysis was performed with LysoTracker Red on p32KO MEFs overexpressing Nmnat3(full) (D) or Nmnat3(v1) (E). One representative experiment out of three shown.
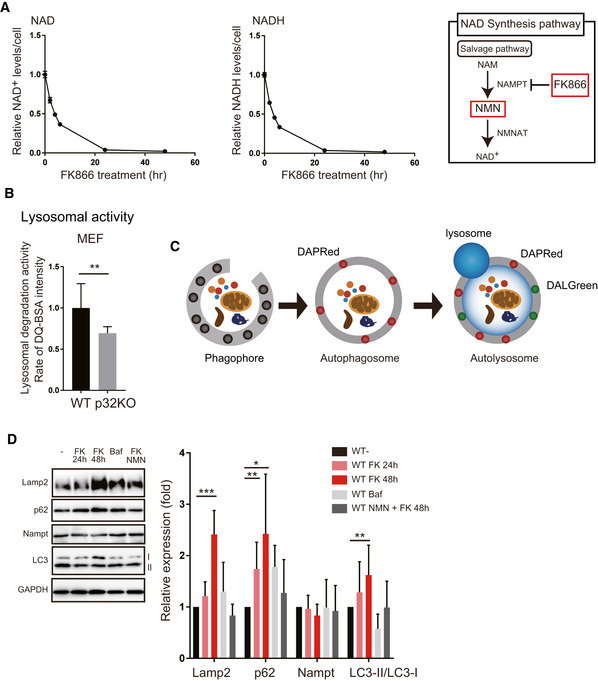
Time course of intracellular NAD+ or NADH content after addition of the Nampt inhibitor, FK866 (n = 3) in WT MEF cells. The right panel shows what FK866 inhibits in the NAD synthesis pathway.
Lysosomal activity was measured by DQ‐BSA in WT and p32KO MEFs. Error bars are presented as mean ± SEM of three independent experiments. Statistical significance was assessed by Student’s t‐test, **P < 0.002.
To monitor autophagosomes and autolysosomes, DAPRed and DALGreen were used. DALGreen fluorescence is enhanced at an acidic pH and is suitable for monitoring the autophagy degradation stage, also known as the autolysosome stage. In contrast, DAPRed has a pH‐independent fluorescence profile and remains fluorescent with almost constant intensity throughout the process of autophagy.
Western blot analysis of Lamp2, p62, Nampt, and LC3. Treatment of WT MEFs with FK866 increased the expression of Lamp2, p62, and LC3. When bafilomycin A or FK866 plus NMN were added, the expression of these proteins did not change. One representative experiment out of three shown. Quantification is shown on the right side. The values are mean ± SD of three independent experiments. Statistical significance was assessed by Student’s t‐test, *P < 0.05, **P < 0.01, ***P < 0.005 (n = 4 per group).
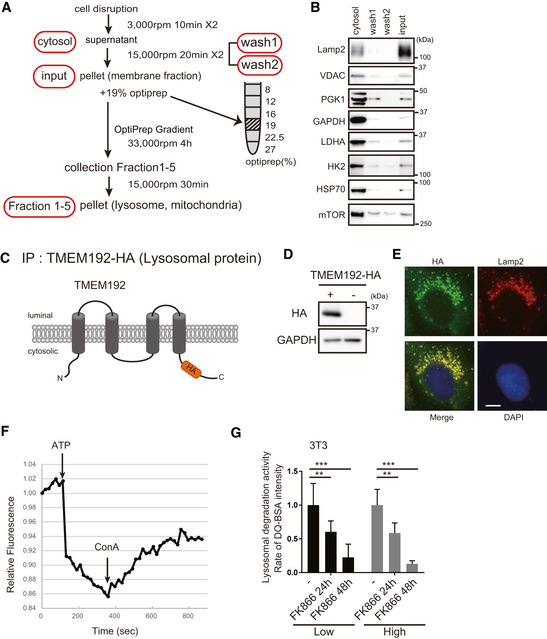
- A
Schema of lysosome purification by centrifugation. The samples were washed twice to reduce cytosolic protein contamination.
- B
The lysosomal fractions in WT MEFs were subjected to Western blotting to examine the presence of characteristic organelle marker proteins. After two washes and centrifugations, the membrane fraction was concentrated, and very little protein was detected, suggesting that this method results in little cytosolic protein contamination.
- C
Schema of the HA‐tagged construct of lysosomal protein TMEM192.
- D, E
The expression of TMEM192 in 3T3‐L1 cells was examined by Western blotting and immunostaining (HA: Green and Lamp2: Red). Scale bar, 10 µm.
- F
Lysosomal fraction isolated with MAG10 was in the active state in 3T3‐L1 cells. V‐ATPase activity was monitored by ACMA, which is a fluorescent pH indicator. The activity was started using ATP and inhibited after addition of concanavalin A (ConA). One representative experiment out of three shown.
- G
Lysosomal activity was measured by DQ‐BSA. Two types of culture medium were used: low‐ and high‐glucose DMEM. Decreased lysosomal proteolytic capacity was observed after FK866 treatment of 3T3‐L1 cells. However, the glucose concentration had no effect. Error bars are presented as mean ± SEM of three independent experiments. Statistical significance was assessed by one‐way ANOVA, **P < 0.005, ***P < 0.001.
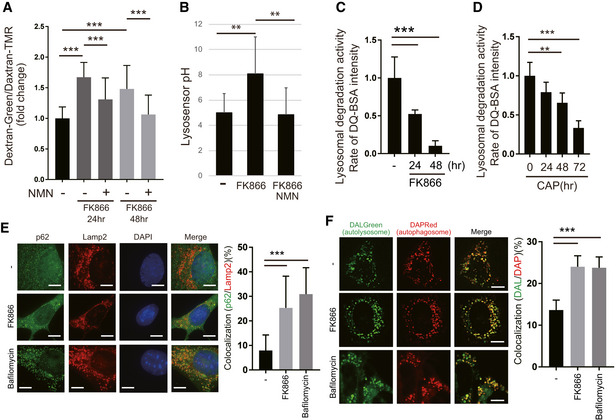
WT MEFs were stained with dextran‐Oregon Green and dextran‐TMRM to examine lysosomal acidification after 10 nM FK866 treatment. FK866 decreased lysosomal acidification, which was rescued by 1 mM NMN pretreatment. The stained MEFs were quantified using a microscope (BZ‐X800, KEYENCE). Error bars are presented as mean ± SEM of three independent experiments. Statistical significance was assessed by one‐way ANOVA, ***P < 0.001.
LysoSensor™ Yellow/Blue DND‐160 was used to detect lysosomal pH function in WT MEFs. Lysosomal acidification was decreased by 10 nM FK866 and rescued by 1 mM NMN. Error bars are presented as mean ± SEM of three independent experiments. Statistical significance was assessed by one‐way ANOVA, **P < 0.01.
Lysosomal proteolytic capacity was decreased in FK866‐treated WT MEFs. DQ‐BSA is bovine serum albumin labeled with a green fluorophore. It is taken up by endocytosis and delivered to the lysosomes. As a monomer, the fluorophore is too concentrated and the signal is quenched. Because DQ‐BSA is degraded by the lysosomal proteases, it releases monomers, which emit fluorescence. The rate of DQ‐BSA hydrolysis (increase in fluorescence units per cell) is a function of lysosomal proteases and was measured using a plate reader over 1 h. The linear range was used to determine the rate. The mean ± SEM rate for each condition is presented in the plot. These values are of a representative experiment. Three independent experiments were performed. Statistical significance was assessed by one‐way ANOVA, ***P < 0.001.
Lysosomal proteolytic capacity was decreased after 1 mM CAP treatment of WT MEFs in a time‐dependent manner, as shown by DQ‐BSA. Error bars are presented as mean ± SEM of three independent experiments. Statistical significance was assessed by one‐way ANOVA, **P < 0.01, ***P < 0.001.
To examine autolysosome formation, we performed immunofluorescence with Lamp2 and p62 antibodies. Lamp2 and p62 were co‐localized after 10 nM FK866 or 50 nM bafilomycin A treatment in WT MEF cells. Scale bars, 10 µm. Quantification is shown on the right. Error bars are presented as mean ± SEM of three independent experiments. Statistical significance was assessed by one‐way ANOVA, ***P < 0.001.
To examine autophagosome and autolysosome formation, we used DAPRed, which indicates autophagosomes, and DALGreen, which indicates autolysosomes. After starvation, co‐localization of both staining agents was observed upon FK866 or bafilomycin A treatment in WT MEF cells. Scale bars, 10 µm. Quantification is shown on the right. Error bars are presented as mean ± SEM of three independent experiments. Statistical significance was assessed by one‐way ANOVA, ***P < 0.001.
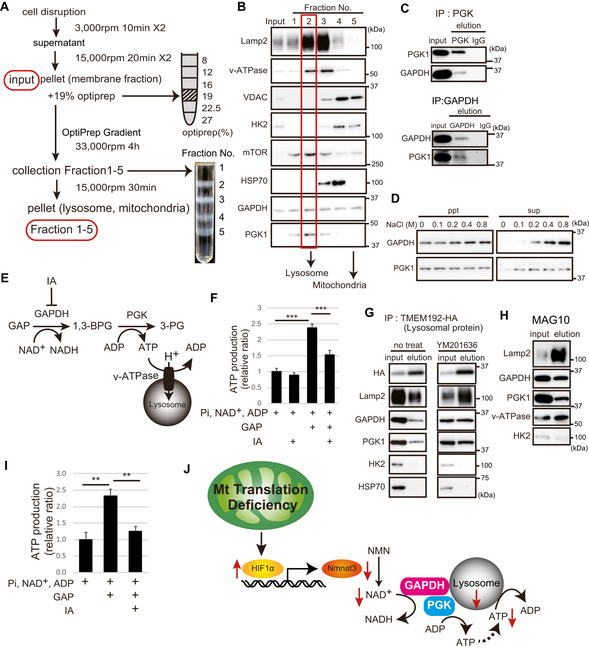
Schema of purification of lysosomes by centrifugation. We applied a low osmotic medium for density gradient centrifugation. After primary organelle enrichment by differential centrifugation from WT MEFs, the subcellular compartments were separated on a discontinuous iodixanol gradient. After membrane pellets were enriched (input), each organelle was purified by an OptiPrep™ density gradient. We separated the five fractions from top to bottom. After ultracentrifugation, five individual fractions became apparent at the individual interphases and were collected, pelleted and used for subsequent analyses.
The Western blots were analyzed for the presence of known organelle marker proteins as indicated. Lysosomal and mitochondrial fractions from WT MEFs were enriched in fractions 2 and 5, respectively.
Lysate of WT MEFs was immunoprecipitated with anti‐GAPDH, anti‐PGK1, and IgG antibodies. Immunoprecipitants were analyzed by Western blotting with anti‐PGK1 and anti‐GAPDH antibody.
After adding various concentrations of NaCl to the lysosomal fraction of WT MEFs, the membrane pellet (bound) and supernatant (free) were subjected to immunoblotting with GAPDH and PGK1 antibodies.
Schematic representation showing that the lysosomal V‐ATPase activity is regulated by the glycolytic enzymes, GAPDH and PGK1. The glycolytic GAPDH activity was measured with GAP as a substrate in the presence of NAD. PGK1 catalyzes the reaction of 1,3‐bisphosphoglycerate (1,3‐BPG) and ADP to form 3‐phosphoglycerate (3‐PG) and ATP.
After addition of GAP, NAD, ADP, and Pi, ATP production in the lysosomal fraction was measured using a luciferin/luciferase kit. Iodoacetate (IA; 4 µM) was used to inhibit GAPDH activity. Three independent experiments were performed. Error bars are presented as mean ± SEM of three independent experiments. Statistical significance was assessed by one‐way ANOVA, ***P < 0.001.
Lysates were prepared from MEF cells expressing Tmem192‐3 × HA [the only enzyme that catalyzes PI(3,5)P2 production in the lysosomal membrane] that were treated with 1 µM YM201636 (PIKfyve inhibitor). The lysosome immunoprecipitation method isolates pure lysosomes with anti‐HA antibody. Immunoblotting of protein markers of various subcellular compartments in purified immunoprecipitated lysosomes are shown.
The lysosomal fraction of WT MEF cells using the Dextran magnetic beads (MAG10) method contained GAPDH and PGK1.
After addition of GAP, NAD, ADP, and Pi, ATP production in the lysosomal fraction isolated by MAG10 was measured using a luciferin/luciferase kit. Iodoacetate (IA; 4 µM) was used to inhibit GAPDH activity. Three independent experiments were performed. Error bars are presented as mean ± SEM of three independent experiments. Statistical significance was assessed by one‐way ANOVA, **P < 0.002.
Schema of this novel study. Mitochondrial translation deficiency suppresses HIF1α‐Nmnat3‐mediated NAD+ production. The glycolytic enzymes, GAPDH and PGK1, are associated with lysosomal vesicles, and NAD+ is required for ATP production around lysosomal vesicles. NAD+ content is essential for lysosomal maintenance.
Similar articles
-
Deficiency of nicotinamide mononucleotide adenylyltransferase 3 (nmnat3) causes hemolytic anemia by altering the glycolytic flow in mature erythrocytes.J Biol Chem. 2014 May 23;289(21):14796-811. doi: 10.1074/jbc.M114.554378. Epub 2014 Apr 16. J Biol Chem. 2014. PMID: 24739386 Free PMC article.
-
Improving lysosomal ferroptosis with NMN administration protects against heart failure.Life Sci Alliance. 2023 Oct 4;6(12):e202302116. doi: 10.26508/lsa.202302116. Print 2023 Dec. Life Sci Alliance. 2023. PMID: 37793777 Free PMC article.
-
Nmnat3 Is Dispensable in Mitochondrial NAD Level Maintenance In Vivo.PLoS One. 2016 Jan 12;11(1):e0147037. doi: 10.1371/journal.pone.0147037. eCollection 2016. PLoS One. 2016. PMID: 26756334 Free PMC article.
-
NAD+ precursor modulates post-ischemic mitochondrial fragmentation and reactive oxygen species generation via SIRT3 dependent mechanisms.Exp Neurol. 2020 Mar;325:113144. doi: 10.1016/j.expneurol.2019.113144. Epub 2019 Dec 16. Exp Neurol. 2020. PMID: 31837320 Free PMC article. Review.
-
Potential Treatment of Lysosomal Storage Disease through Modulation of the Mitochondrial-Lysosomal Axis.Cells. 2021 Feb 17;10(2):420. doi: 10.3390/cells10020420. Cells. 2021. PMID: 33671306 Free PMC article. Review.
Cited by
-
Can niacin supplementation prevent congenital malformations associated with maternal use of proton pump inhibitors?Eur J Nutr. 2023 Mar;62(2):1051-1053. doi: 10.1007/s00394-022-03060-1. Epub 2022 Nov 28. Eur J Nutr. 2023. PMID: 36441236 No abstract available.
-
Glycolysis-Mediated Activation of v-ATPase by Nicotinamide Mononucleotide Ameliorates Lipid-Induced Cardiomyopathy by Repressing the CD36-TLR4 Axis.Circ Res. 2024 Mar;134(5):505-525. doi: 10.1161/CIRCRESAHA.123.322910. Epub 2024 Feb 7. Circ Res. 2024. PMID: 38422177 Free PMC article.
-
Role of Lysosomal Acidification Dysfunction in Mesenchymal Stem Cell Senescence.Front Cell Dev Biol. 2022 Feb 7;10:817877. doi: 10.3389/fcell.2022.817877. eCollection 2022. Front Cell Dev Biol. 2022. PMID: 35198560 Free PMC article. Review.
-
Activation of the integrated stress response confers vulnerability to mitoribosome-targeting antibiotics in melanoma.J Exp Med. 2021 Sep 6;218(9):e20210571. doi: 10.1084/jem.20210571. Epub 2021 Jul 21. J Exp Med. 2021. PMID: 34287642 Free PMC article.
-
Lysosomal dysfunction and overload of nucleosides in thymidine phosphorylase deficiency of MNGIE.J Transl Med. 2024 May 13;22(1):449. doi: 10.1186/s12967-024-05275-8. J Transl Med. 2024. PMID: 38741129 Free PMC article.
References
-
- Bissig C, Hurbain I, Raposo G, van Niel G (2017) PIKfyve activity regulates reformation of terminal storage lysosomes from endolysosomes. Traffic 18: 747–757 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
