

The Axenfeld-Rieger Syndrome Gene FOXC1 Contributes to Left-Right Patterning
- PMID: 33530637
- PMCID: PMC7912076
- DOI: 10.3390/genes12020170
The Axenfeld-Rieger Syndrome Gene FOXC1 Contributes to Left-Right Patterning
Abstract
Precise spatiotemporal expression of the Nodal-Lefty-Pitx2 cascade in the lateral plate mesoderm establishes the left-right axis, which provides vital cues for correct organ formation and function. Mutations of one cascade constituent PITX2 and, separately, the Forkhead transcription factor FOXC1 independently cause a multi-system disorder known as Axenfeld-Rieger syndrome (ARS). Since cardiac involvement is an established ARS phenotype and because disrupted left-right patterning can cause congenital heart defects, we investigated in zebrafish whether foxc1 contributes to organ laterality or situs. We demonstrate that CRISPR/Cas9-generated foxc1a and foxc1b mutants exhibit abnormal cardiac looping and that the prevalence of cardiac situs defects is increased in foxc1a-/-; foxc1b-/- homozygotes. Similarly, double homozygotes exhibit isomerism of the liver and pancreas, which are key features of abnormal gut situs. Placement of the asymmetric visceral organs relative to the midline was also perturbed by mRNA overexpression of foxc1a and foxc1b. In addition, an analysis of the left-right patterning components, identified in the lateral plate mesoderm of foxc1 mutants, reduced or abolished the expression of the NODAL antagonist lefty2. Together, these data reveal a novel contribution from foxc1 to left-right patterning, demonstrating that this role is sensitive to foxc1 gene dosage, and provide a plausible mechanism for the incidence of congenital heart defects in Axenfeld-Rieger syndrome patients.
Keywords: Axenfeld–Rieger syndrome; FOXC1; LEFTY; left–right patterning; zebrafish.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
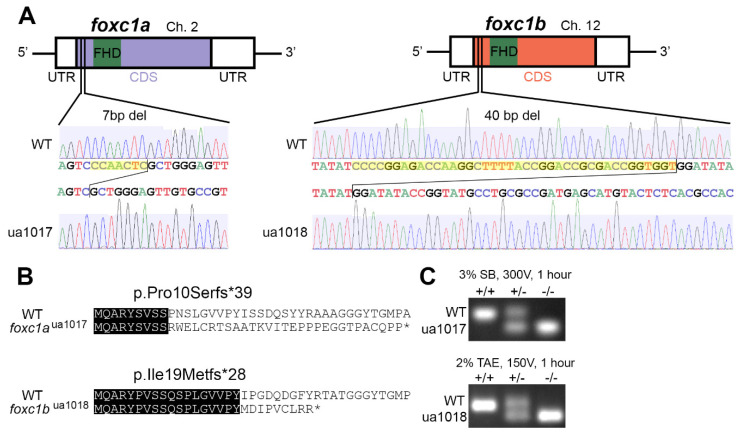
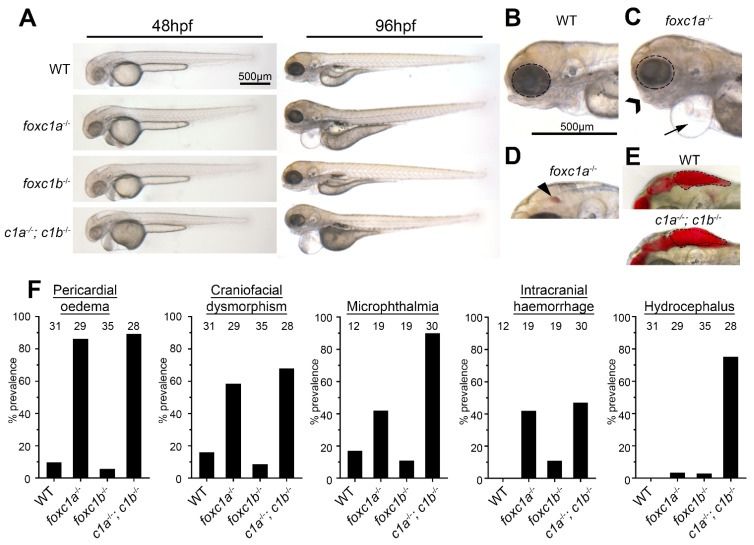
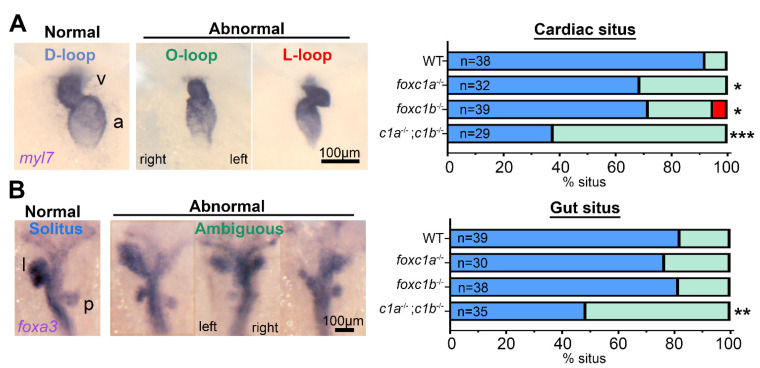
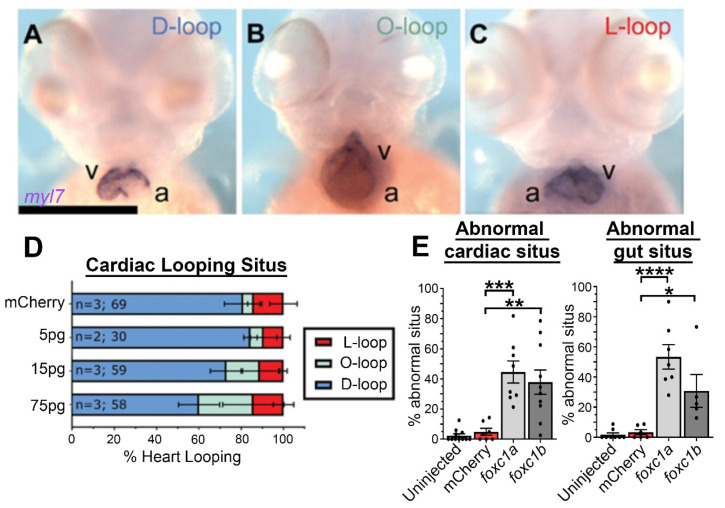
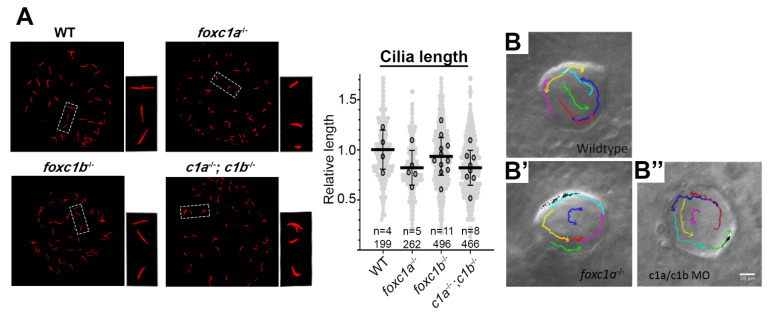
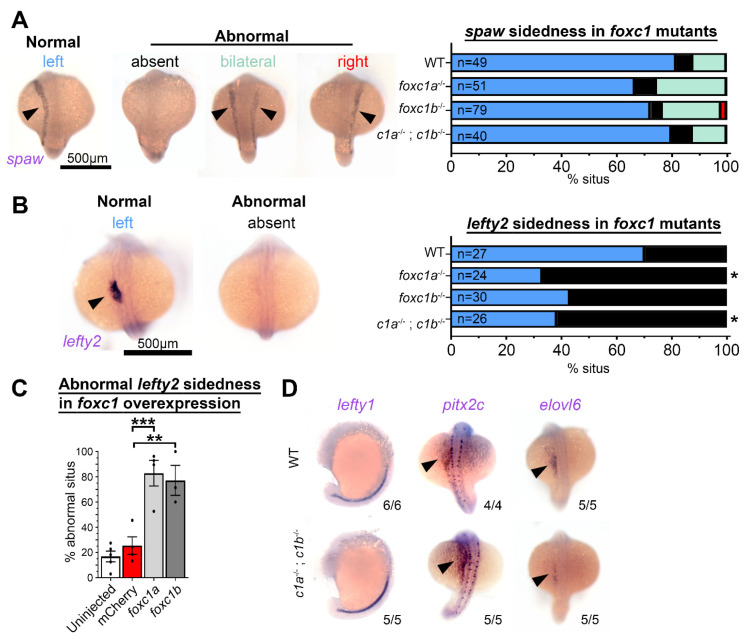
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
