

The Infantile Leukoencephalopathy-Associated Mutation of C11ORF73/HIKESHI Proteins Generates de novo Interactive Activity with Filamin A, Inhibiting Oligodendroglial Cell Morphological Differentiation
- PMID: 33535532
- PMCID: PMC7912763
- DOI: 10.3390/medicines8020009
The Infantile Leukoencephalopathy-Associated Mutation of C11ORF73/HIKESHI Proteins Generates de novo Interactive Activity with Filamin A, Inhibiting Oligodendroglial Cell Morphological Differentiation
Abstract
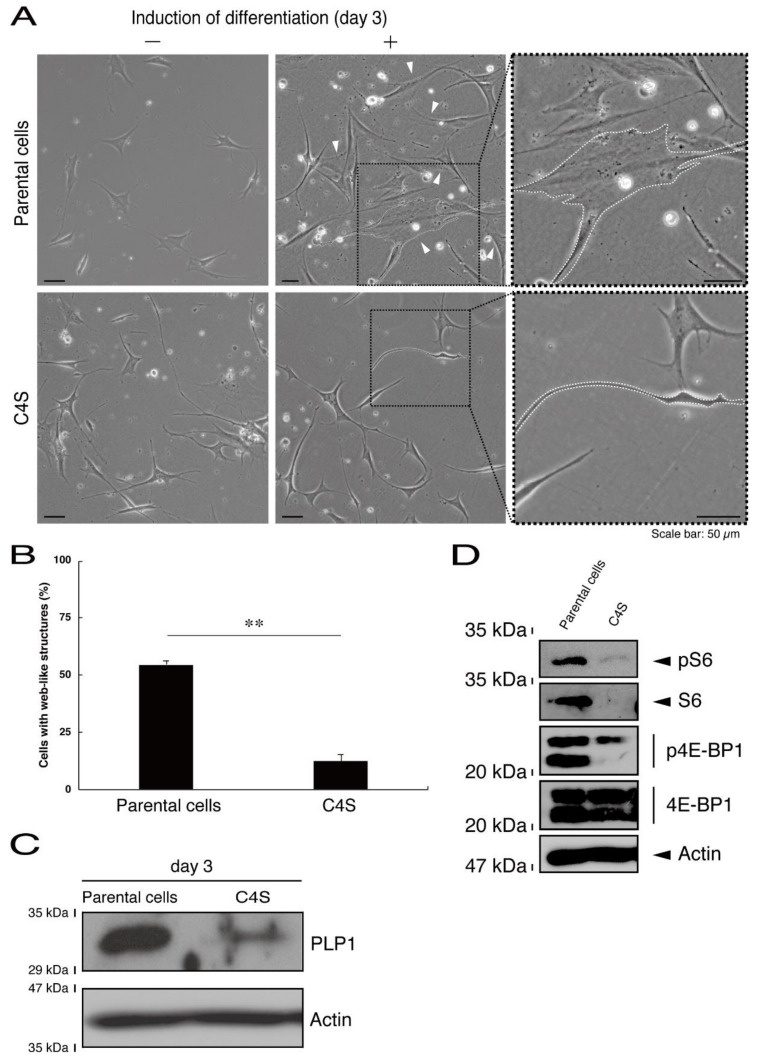
Genetic hypomyelinating diseases are a heterogeneous group of disorders involving the white matter. One infantile hypomyelinating leukoencephalopathy is associated with the homozygous variant (Cys4-to-Ser (C4S)) of the c11orf73 gene. Methods: We observed that in mouse oligodendroglial FBD-102b cells, the C4S mutant proteins but not the wild type ones of C11orf73 are microscopically localized in the lysosome. And, they downregulate lysosome-related signaling in an immunoblotting technique. Results: The C4S mutant proteins specifically interact with Filamin A, which is known to anchor transmembrane proteins to the actin cytoskeleton; the C4S mutant proteins and Filamin A are also observed in the lysosome fraction. While parental FBD-102b cells and cells harboring the wild type constructs exhibit morphological differentiation, cells harboring C4S mutant constructs do not. It may be that morphological differentiation is inhibited because expression of these C4S mutant proteins leads to defects in the actin cytoskeletal network involving Filamin A. Conclusions: The findings that leukoencephalopathy-associated C11ORF73 mutant proteins specifically interact with Filamin A, are localized in the lysosome, and inhibit morphological differentiation shed light on the molecular and cellular pathological mechanisms that underlie infantile hypomyelinating leukoencephalopathy.
Keywords: C11orf73; Filamin A; HLD13; leukoencephalopathy; oligodendrocyte differentiation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures









References
LinkOut - more resources
Full Text Sources
Other Literature Sources
