

Recurrent deletions in the SARS-CoV-2 spike glycoprotein drive antibody escape
- PMID: 33536258
- PMCID: PMC7971772
- DOI: 10.1126/science.abf6950
Recurrent deletions in the SARS-CoV-2 spike glycoprotein drive antibody escape
Abstract
Zoonotic pandemics, such as that caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), can follow the spillover of animal viruses into highly susceptible human populations. The descendants of these viruses have adapted to the human host and evolved to evade immune pressure. Coronaviruses acquire substitutions more slowly than other RNA viruses. In the spike glycoprotein, we found that recurrent deletions overcome this slow substitution rate. Deletion variants arise in diverse genetic and geographic backgrounds, transmit efficiently, and are present in novel lineages, including those of current global concern. They frequently occupy recurrent deletion regions (RDRs), which map to defined antibody epitopes. Deletions in RDRs confer resistance to neutralizing antibodies. By altering stretches of amino acids, deletions appear to accelerate SARS-CoV-2 antigenic evolution and may, more generally, drive adaptive evolution.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Figures
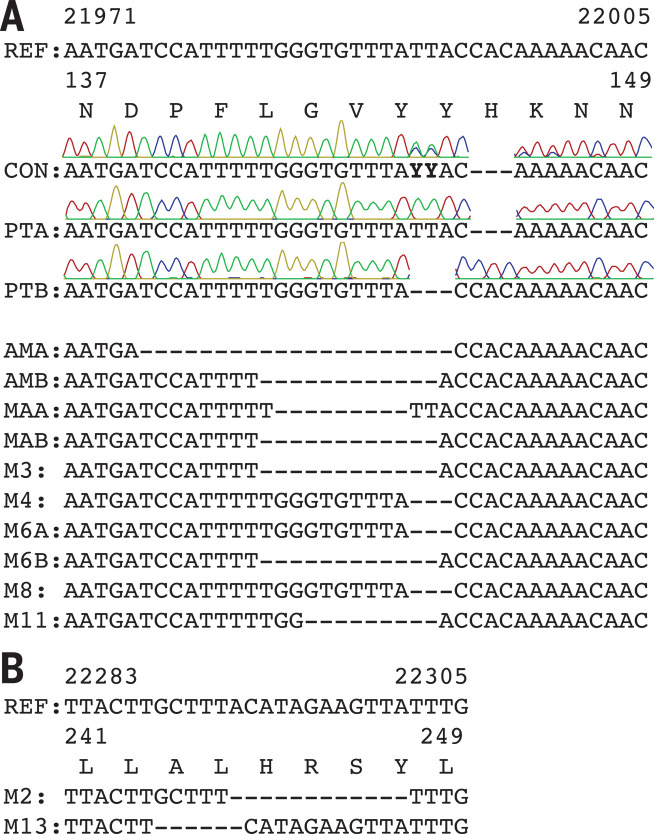
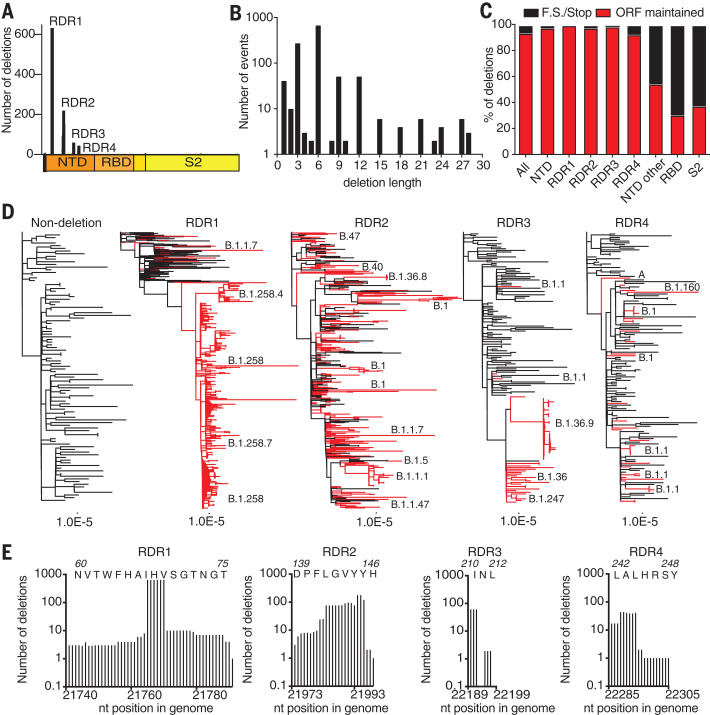
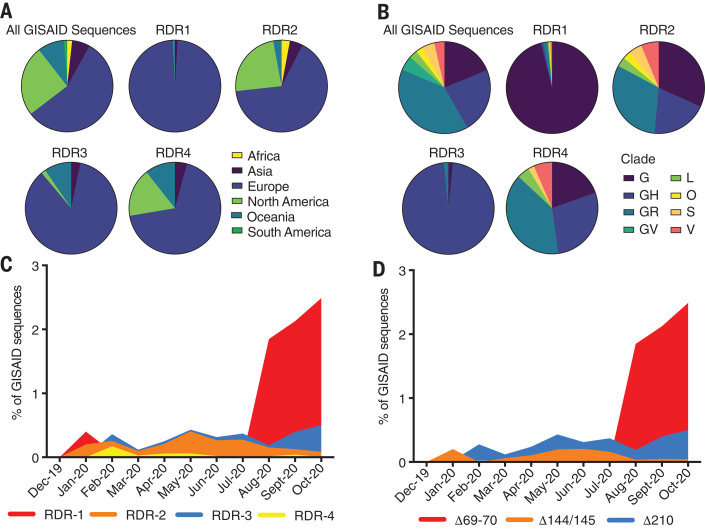
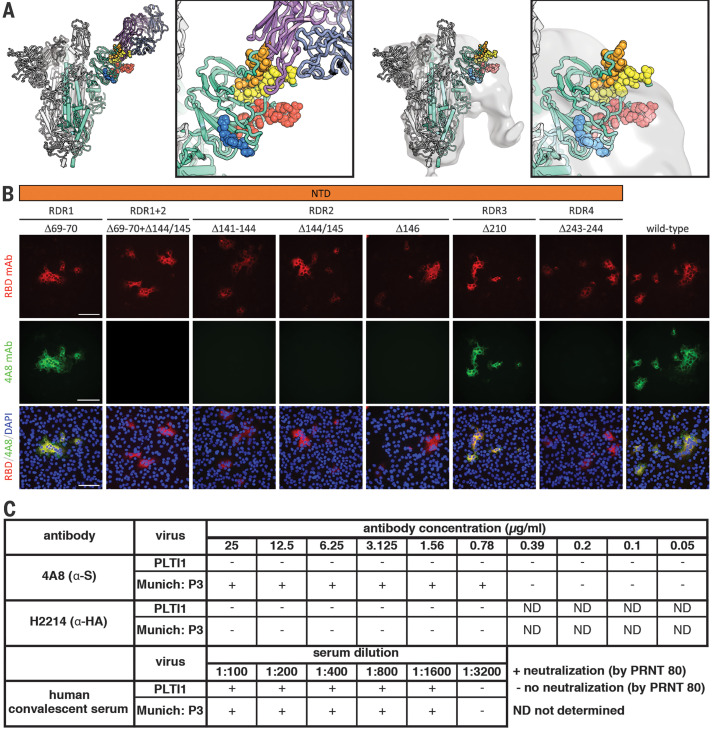
Comment in
-
Adapt or perish: SARS-CoV-2 antibody escape variants defined by deletions in the Spike N-terminal Domain.Signal Transduct Target Ther. 2021 Apr 24;6(1):164. doi: 10.1038/s41392-021-00601-8. Signal Transduct Target Ther. 2021. PMID: 33895775 Free PMC article. No abstract available.
-
COVID-19 variants as moving targets and how to stop them by glycoengineered whole-virus vaccines.Virulence. 2021 Dec;12(1):1717-1720. doi: 10.1080/21505594.2021.1939924. Virulence. 2021. PMID: 34304693 Free PMC article. No abstract available.
References
-
- Wu F., Zhao S., Yu B., Chen Y.-M., Wang W., Song Z.-G., Hu Y., Tao Z.-W., Tian J.-H., Pei Y.-Y., Yuan M.-L., Zhang Y.-L., Dai F.-H., Liu Y., Wang Q.-M., Zheng J.-J., Xu L., Holmes E. C., Zhang Y.-Z., A new coronavirus associated with human respiratory disease in China. Nature 579, 265–269 (2020). 10.1038/s41586-020-2008-3 - DOI - PMC - PubMed
-
- Zhou H., Chen X., Hu T., Li J., Song H., Liu Y., Wang P., Liu D., Yang J., Holmes E. C., Hughes A. C., Bi Y., Shi W., A Novel Bat Coronavirus Closely Related to SARS-CoV-2 Contains Natural Insertions at the S1/S2 Cleavage Site of the Spike Protein. Curr. Biol. 30, 3896 (2020). 10.1016/j.cub.2020.09.030 - DOI - PMC - PubMed
-
- Lam T. T., Jia N., Zhang Y.-W., Shum M. H.-H., Jiang J.-F., Zhu H.-C., Tong Y.-G., Shi Y.-X., Ni X.-B., Liao Y.-S., Li W.-J., Jiang B.-G., Wei W., Yuan T.-T., Zheng K., Cui X.-M., Li J., Pei G.-Q., Qiang X., Cheung W. Y.-M., Li L.-F., Sun F.-F., Qin S., Huang J.-C., Leung G. M., Holmes E. C., Hu Y.-L., Guan Y., Cao W.-C., Identifying SARS-CoV-2-related coronaviruses in Malayan pangolins. Nature 583, 282–285 (2020). 10.1038/s41586-020-2169-0 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
