

Macrophages inhibit and enhance endometriosis depending on their origin
- PMID: 33536334
- PMCID: PMC8017702
- DOI: 10.1073/pnas.2013776118
Macrophages inhibit and enhance endometriosis depending on their origin
Abstract
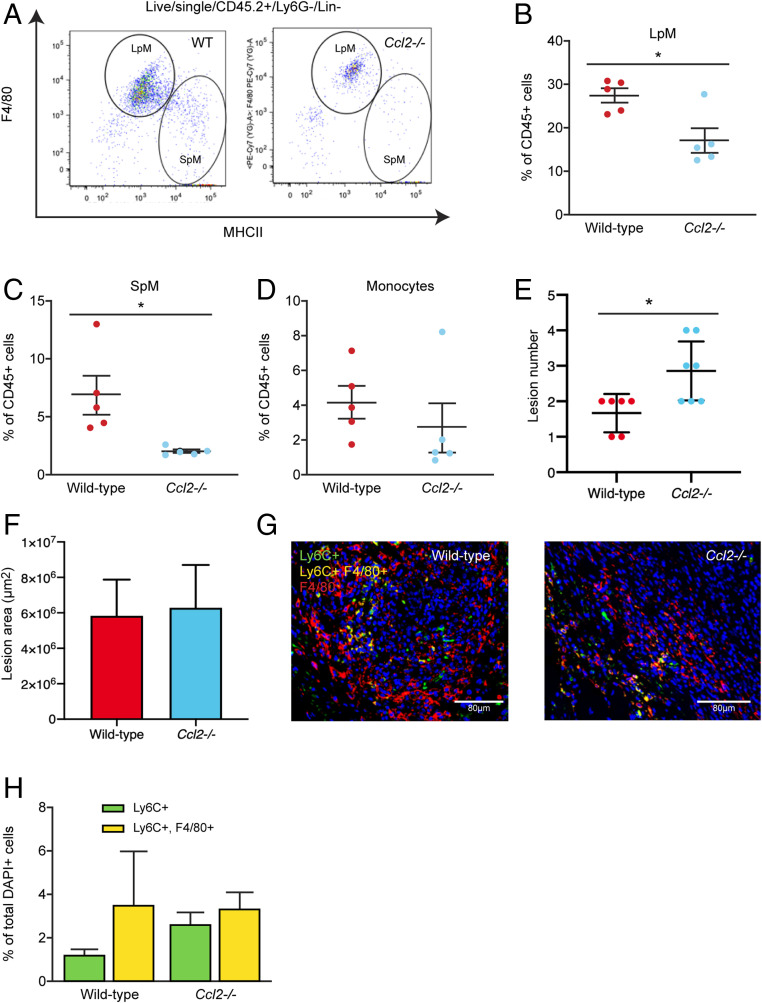
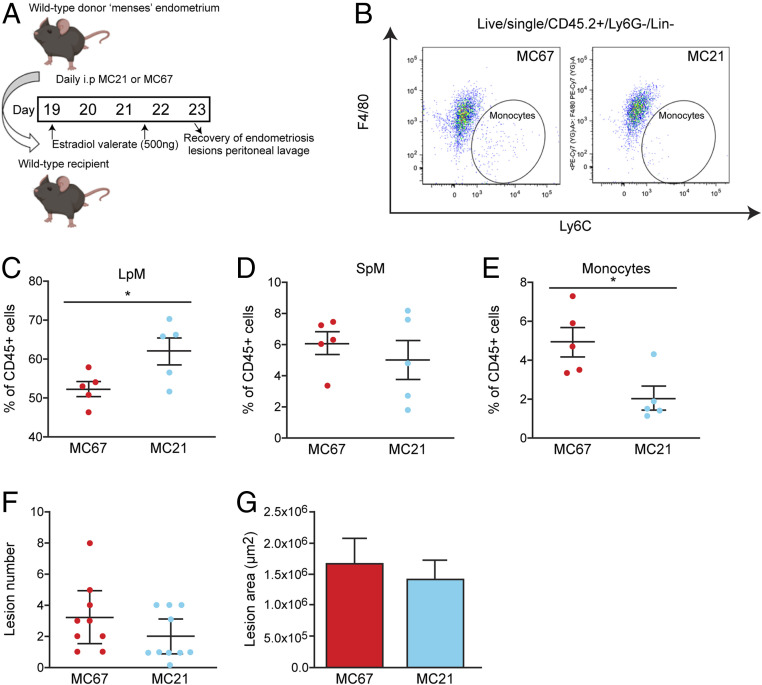
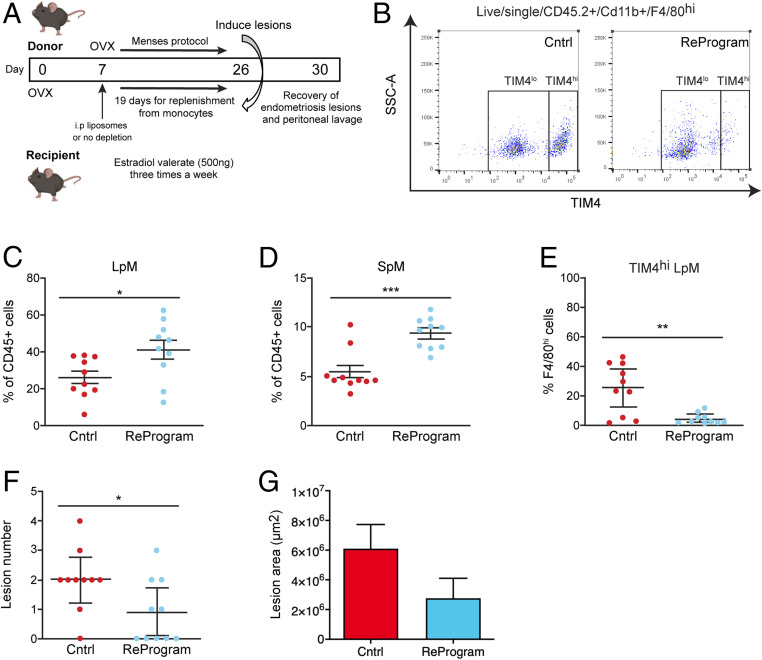
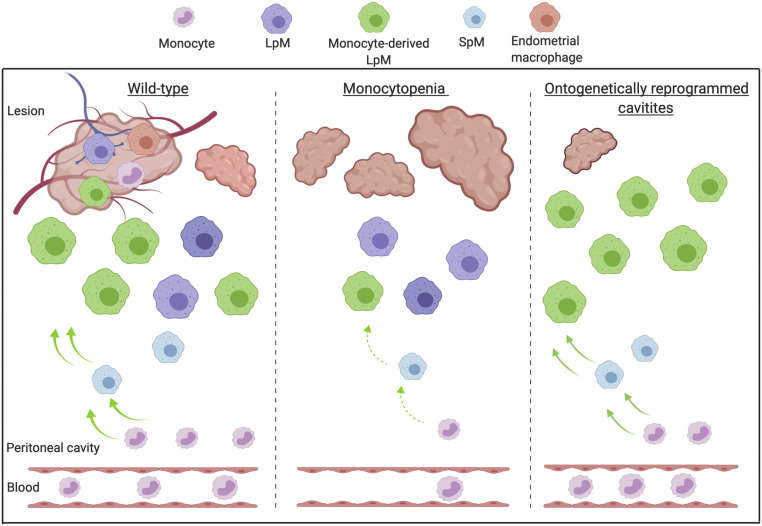
Macrophages are intimately involved in the pathophysiology of endometriosis, a chronic inflammatory disorder characterized by the growth of endometrial-like tissue (lesions) outside the uterus. By combining genetic and pharmacological monocyte and macrophage depletion strategies we determined the ontogeny and function of macrophages in a mouse model of induced endometriosis. We demonstrate that lesion-resident macrophages are derived from eutopic endometrial tissue, infiltrating large peritoneal macrophages (LpM) and monocytes. Furthermore, we found endometriosis to trigger continuous recruitment of monocytes and expansion of CCR2+ LpM. Depletion of eutopic endometrial macrophages results in smaller endometriosis lesions, whereas constitutive inhibition of monocyte recruitment significantly reduces peritoneal macrophage populations and increases the number of lesions. Reprogramming the ontogeny of peritoneal macrophages such that embryo-derived LpM are replaced by monocyte-derived LpM decreases the number of lesions that develop. We propose a putative model whereby endometrial macrophages are "proendometriosis" while newly recruited monocyte-derived macrophages, possibly in LpM form, are "antiendometriosis." These observations highlight the importance of monocyte-derived macrophages in limiting disease progression.
Keywords: lesion; ontogeny; phenotype.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
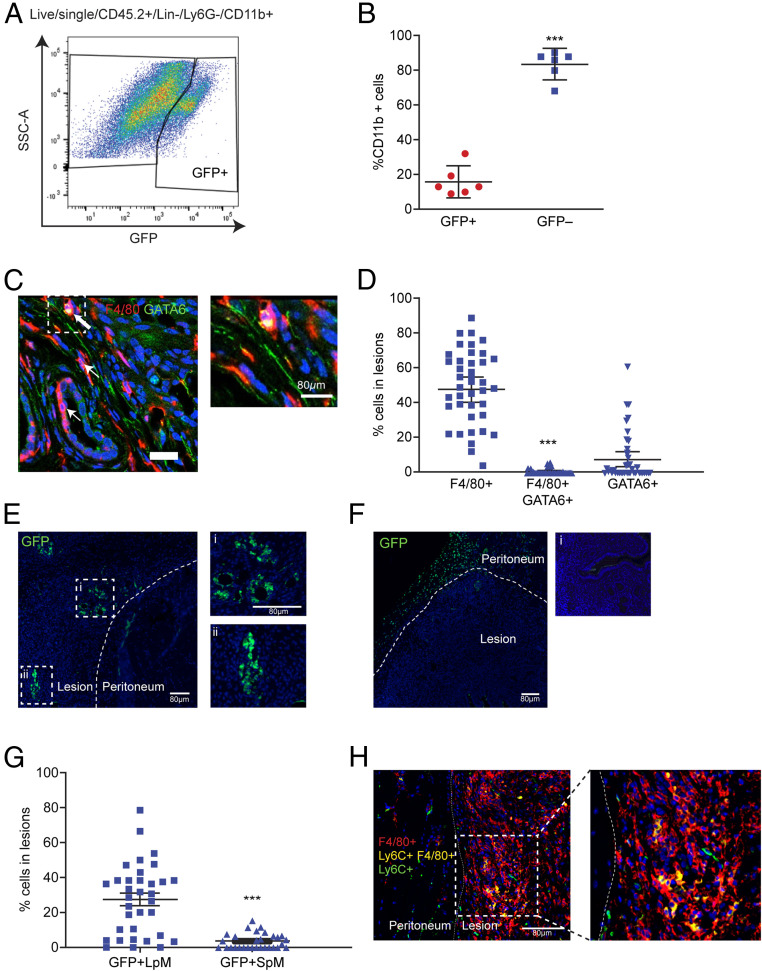
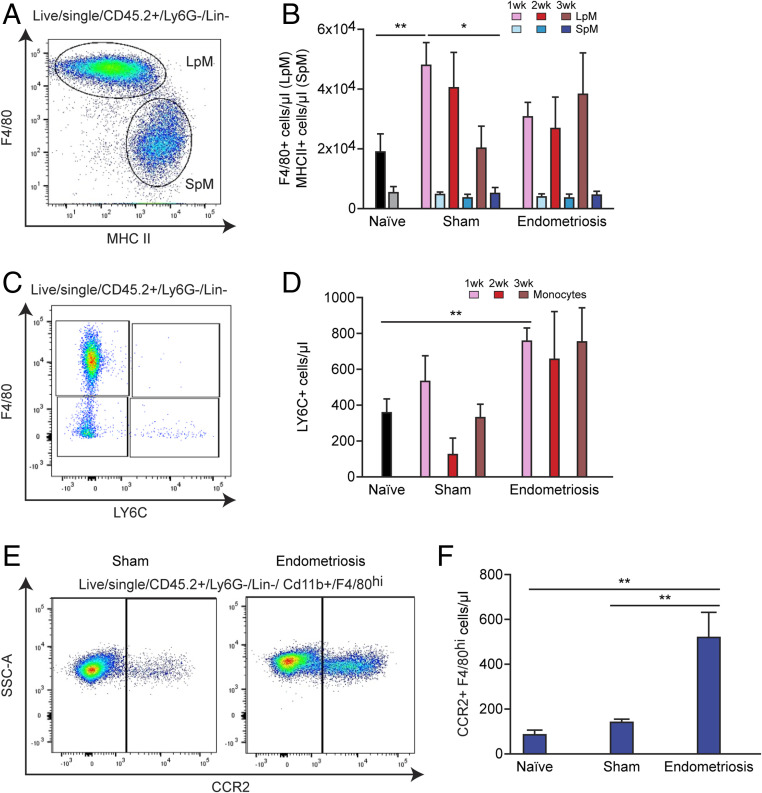
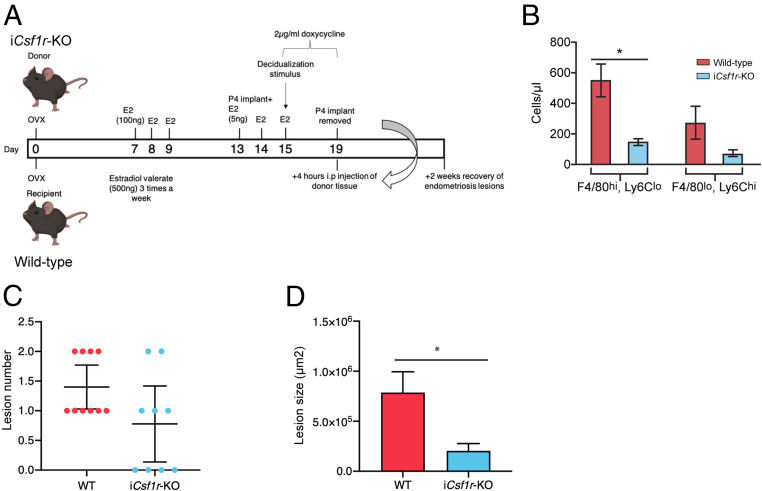
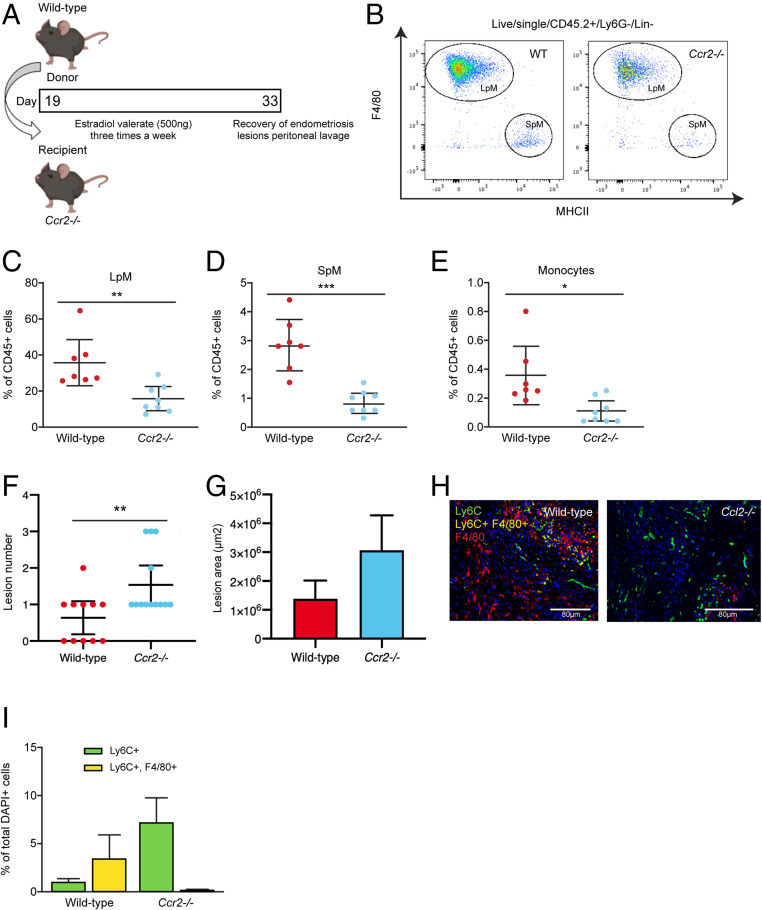
References
-
- Ginhoux F., Jung S., Monocytes and macrophages: Developmental pathways and tissue homeostasis. Nat. Rev. Immunol. 14, 392–404 (2014). - PubMed
-
- Ginhoux F., Schultze J. L., Murray P. J., Ochando J., Biswas S. K., New insights into the multidimensional concept of macrophage ontogeny, activation and function. Nat. Immunol. 17, 34–40 (2016). - PubMed
-
- Schulz C., et al. ., A lineage of myeloid cells independent of Myb and hematopoietic stem cells. Science 336, 86–90 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
