

Mitochondrial Dysfunction in Astrocytes: A Role in Parkinson's Disease?
- PMID: 33537300
- PMCID: PMC7849831
- DOI: 10.3389/fcell.2020.608026
Mitochondrial Dysfunction in Astrocytes: A Role in Parkinson's Disease?
Abstract
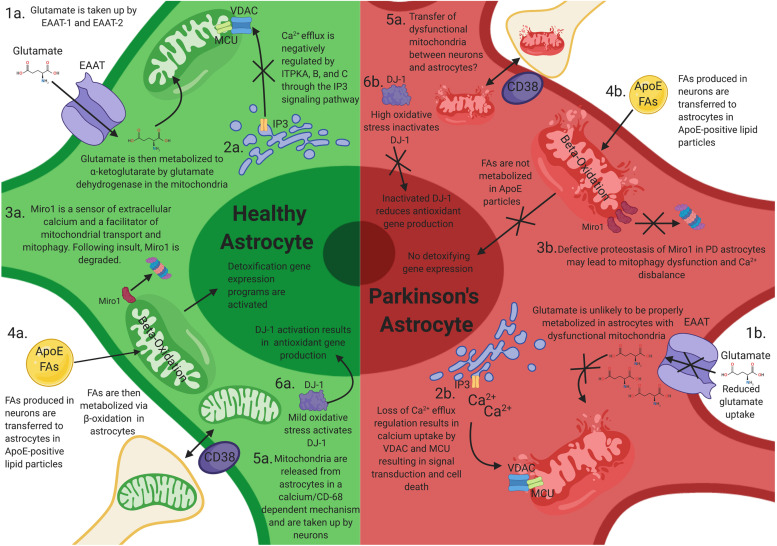
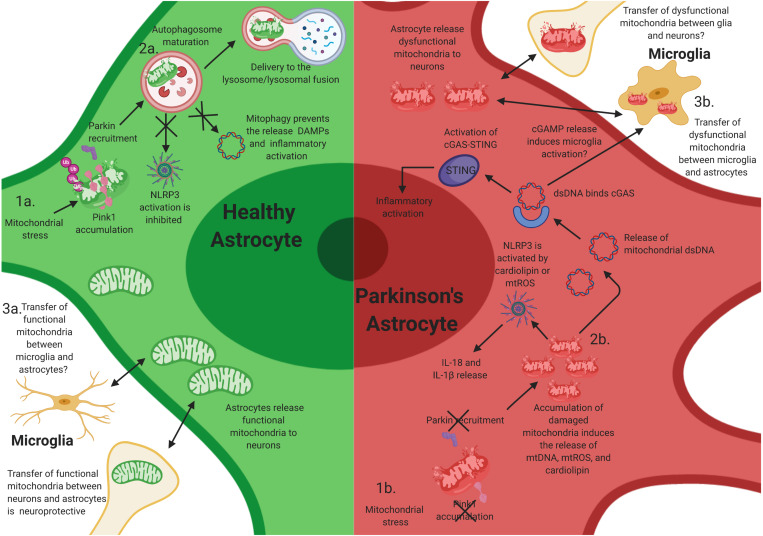
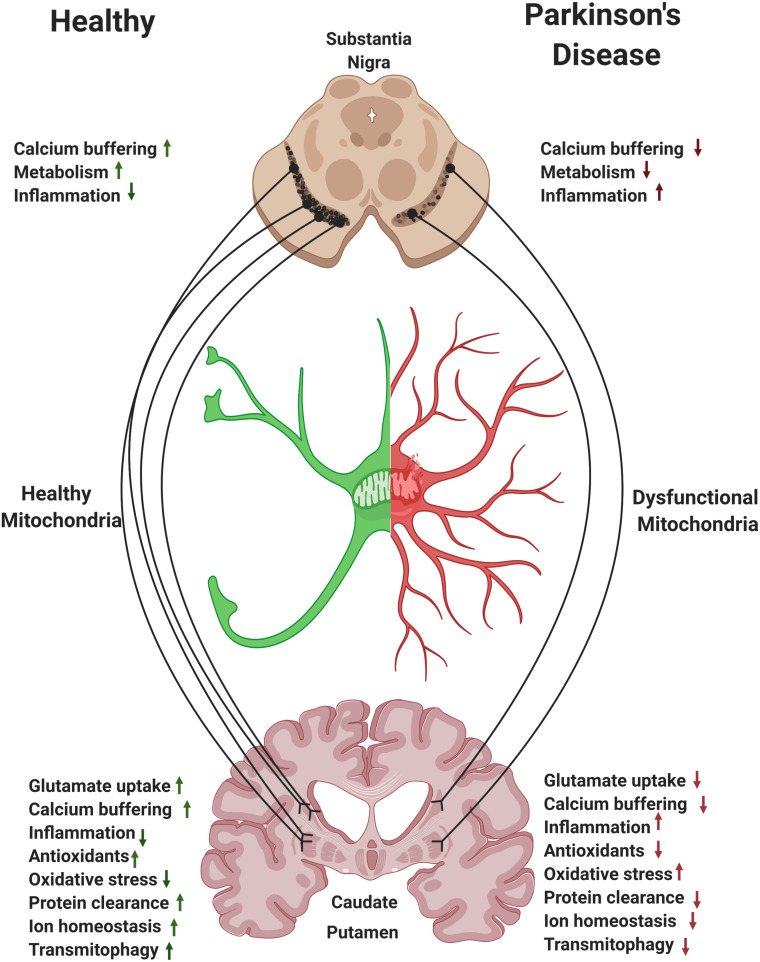
Mitochondrial dysfunction is a hallmark of Parkinson's disease (PD). Astrocytes are the most abundant glial cell type in the brain and are thought to play a pivotal role in the progression of PD. Emerging evidence suggests that many astrocytic functions, including glutamate metabolism, Ca2+ signaling, fatty acid metabolism, antioxidant production, and inflammation are dependent on healthy mitochondria. Here, we review how mitochondrial dysfunction impacts astrocytes, highlighting translational gaps and opening new questions for therapeutic development.
Keywords: NLRP3; PINK1/Parkin pathway; Parkinson’s disease; astrocyte; cGAS/STING pathway; inflammation; mitochondria.
Copyright © 2021 Bantle, Hirst, Weihofen and Shlevkov.
Conflict of interest statement
All authors are employees and shareholders of Biogen.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous