

Completion of neuronal remodeling prompts myelination along developing motor axon branches
- PMID: 33538762
- PMCID: PMC7868780
- DOI: 10.1083/jcb.201911114
Completion of neuronal remodeling prompts myelination along developing motor axon branches
Abstract
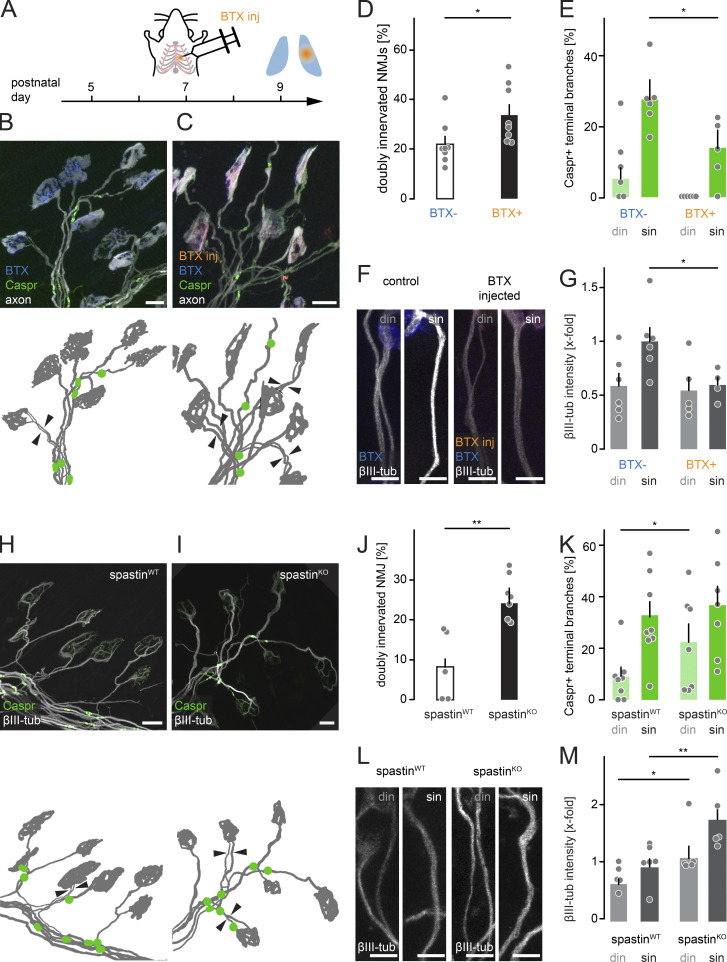
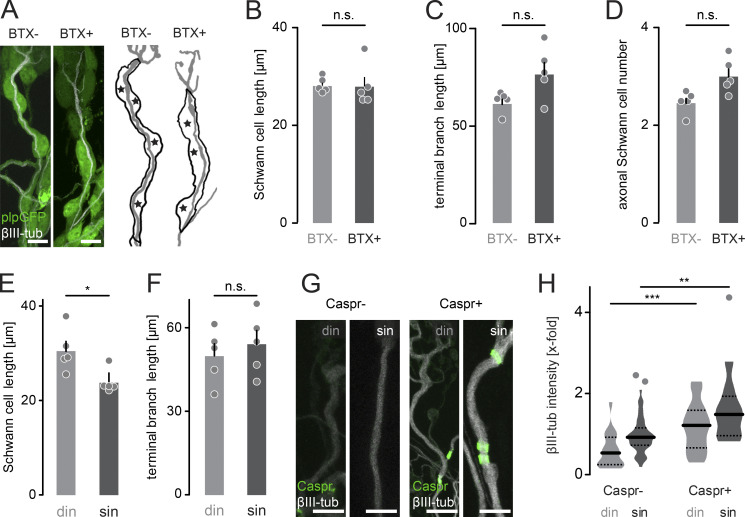
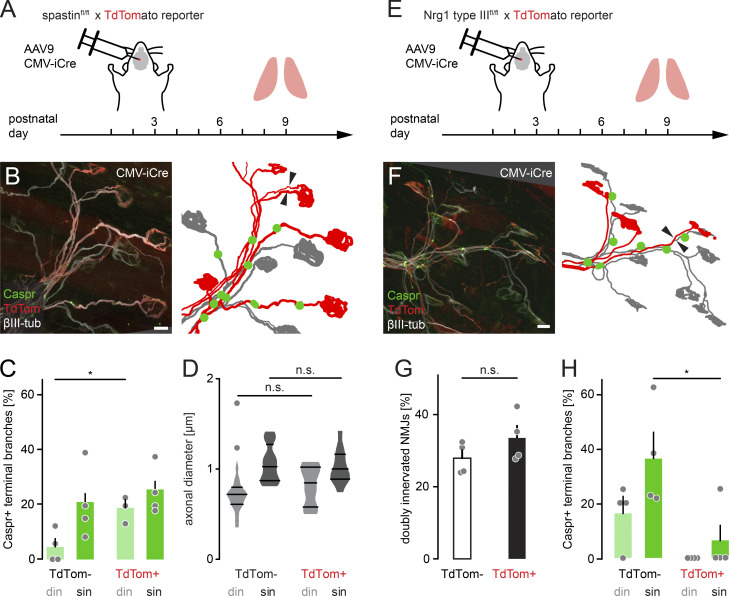
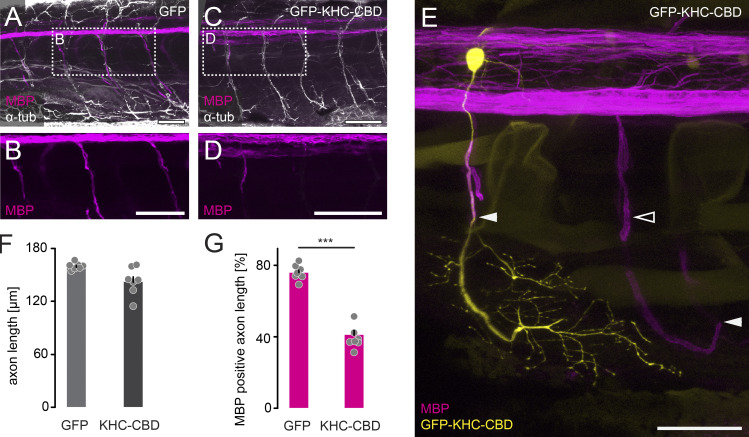
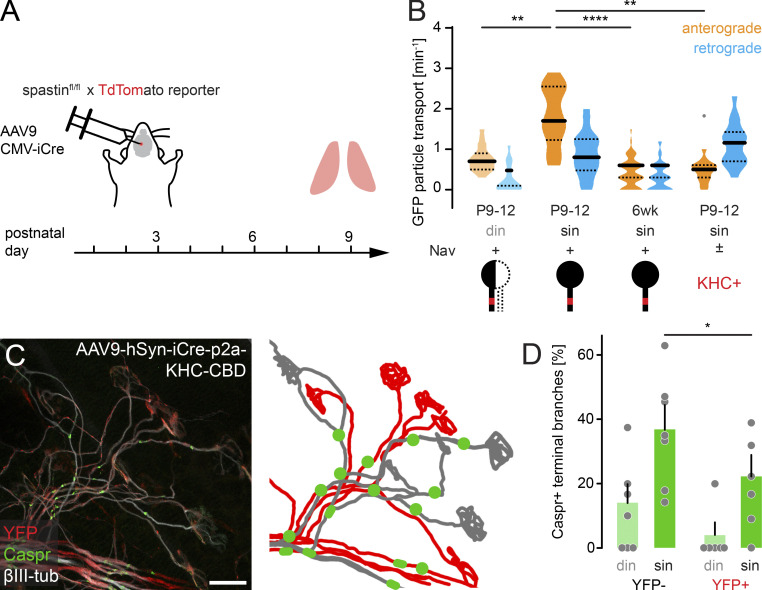
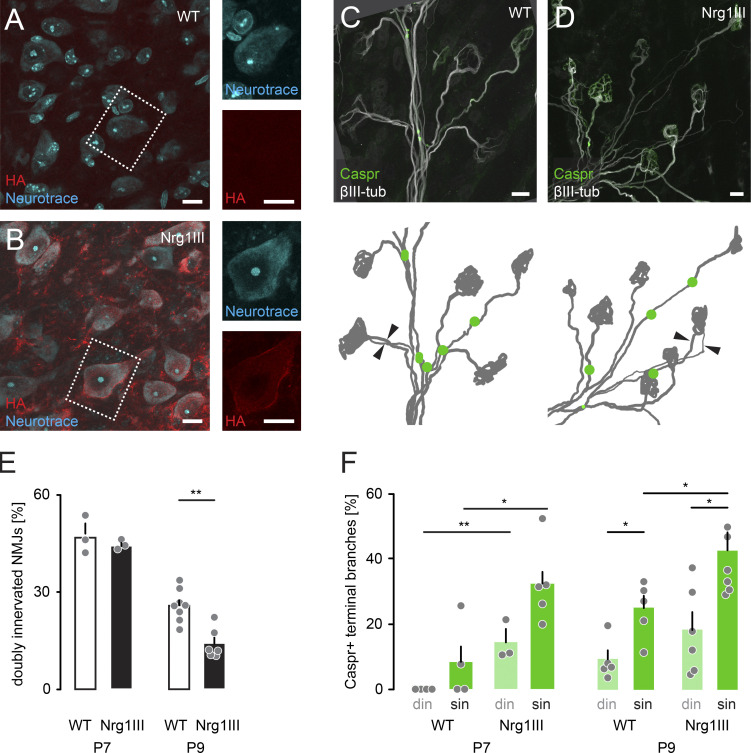
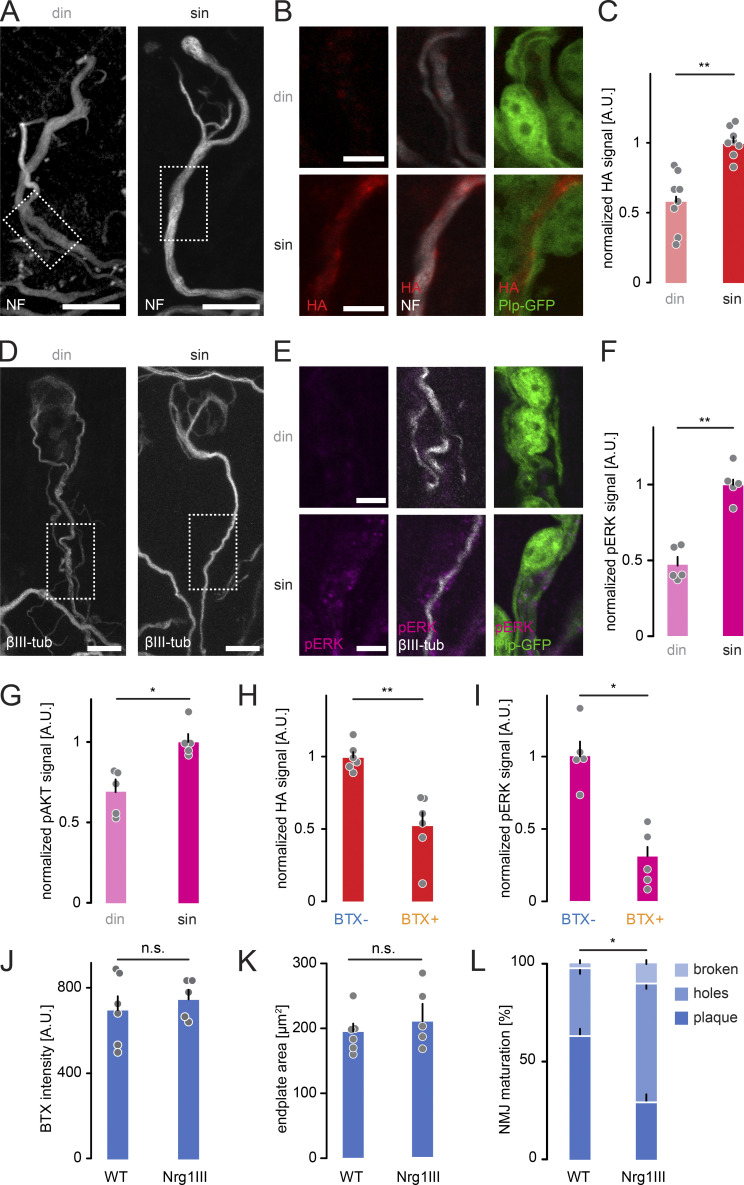
Neuronal remodeling and myelination are two fundamental processes during neurodevelopment. How they influence each other remains largely unknown, even though their coordinated execution is critical for circuit function and often disrupted in neuropsychiatric disorders. It is unclear whether myelination stabilizes axon branches during remodeling or whether ongoing remodeling delays myelination. By modulating synaptic transmission, cytoskeletal dynamics, and axonal transport in mouse motor axons, we show that local axon remodeling delays myelination onset and node formation. Conversely, glial differentiation does not determine the outcome of axon remodeling. Delayed myelination is not due to a limited supply of structural components of the axon-glial unit but rather is triggered by increased transport of signaling factors that initiate myelination, such as neuregulin. Further, transport of promyelinating signals is regulated via local cytoskeletal maturation related to activity-dependent competition. Our study reveals an axon branch-specific fine-tuning mechanism that locally coordinates axon remodeling and myelination.
© 2021 Wang et al.
Figures
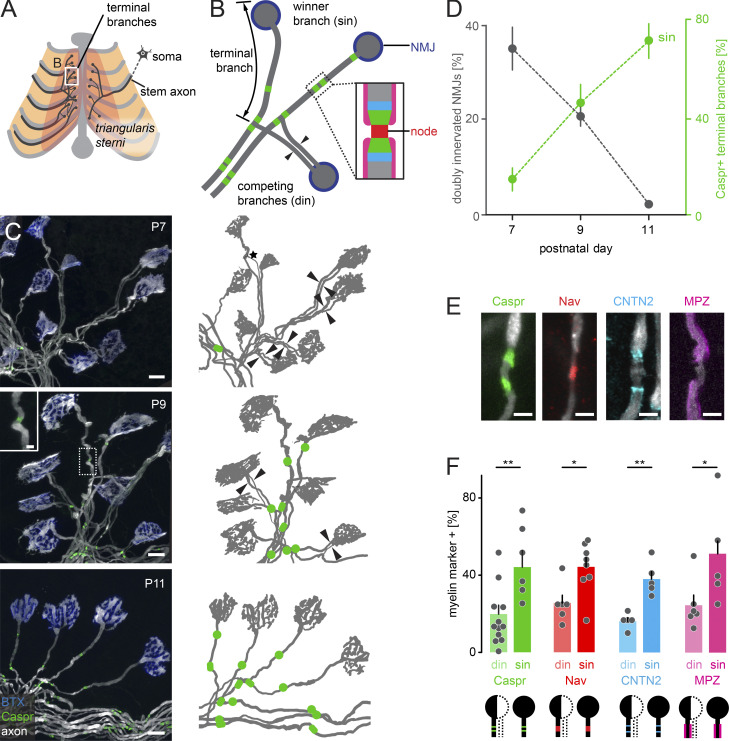
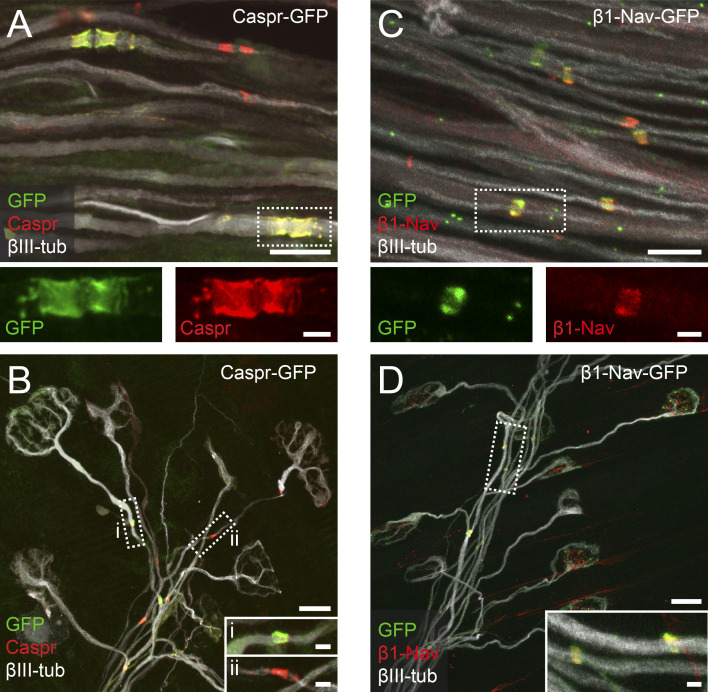
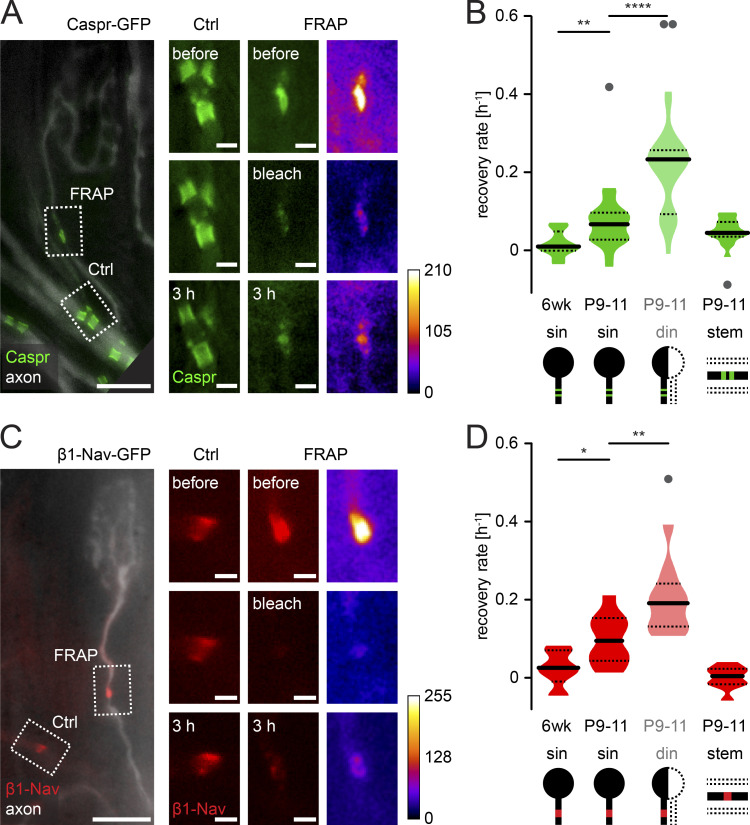
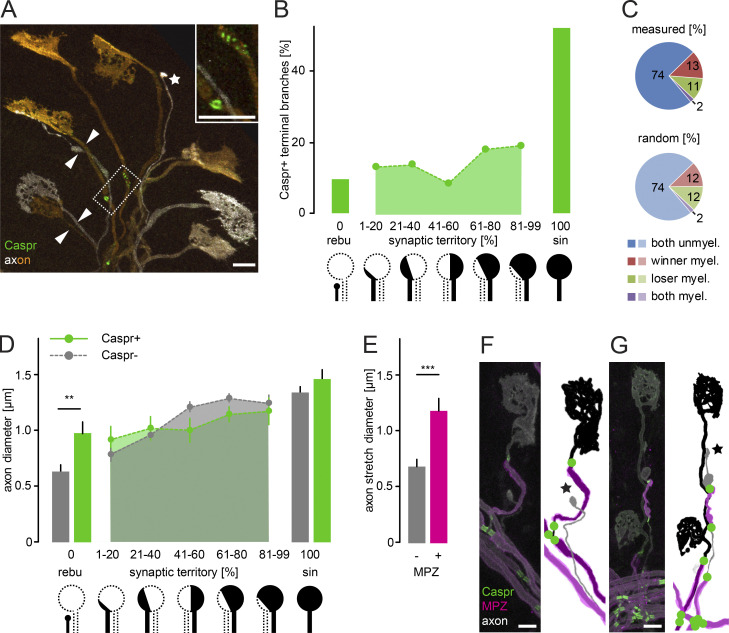
Comment in
-
Lose it to use it.J Cell Biol. 2021 Apr 5;220(4):e202102030. doi: 10.1083/jcb.202102030. J Cell Biol. 2021. PMID: 33734302 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
