

Gating the pore of the calcium-activated chloride channel TMEM16A
- PMID: 33542223
- PMCID: PMC7862301
- DOI: 10.1038/s41467-020-20787-9
Gating the pore of the calcium-activated chloride channel TMEM16A
Abstract
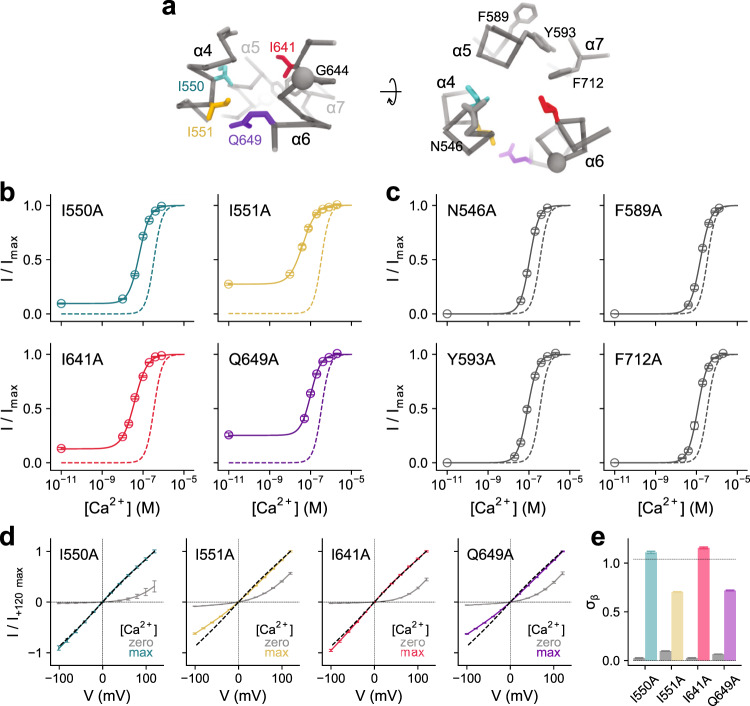
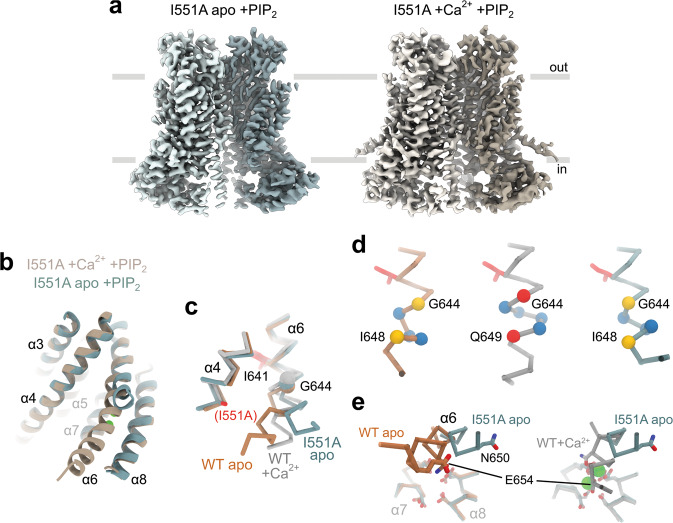
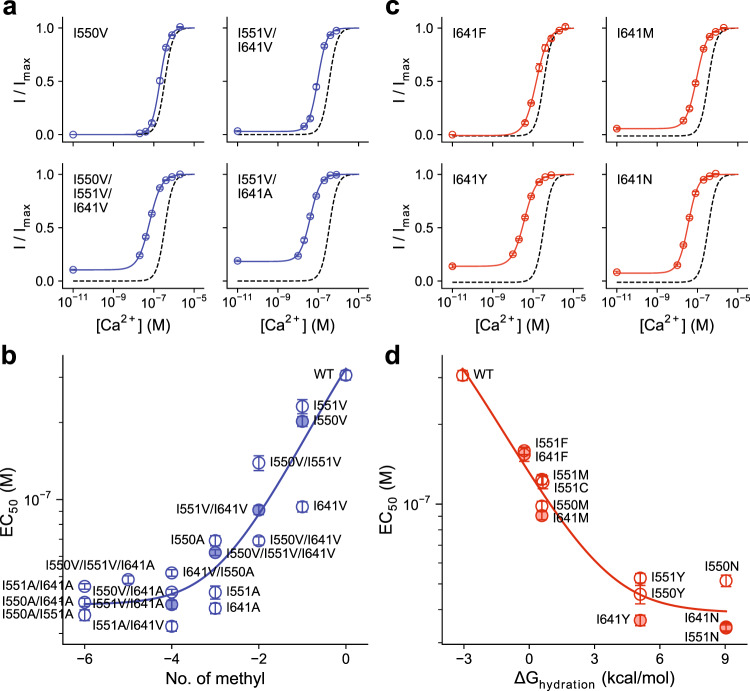
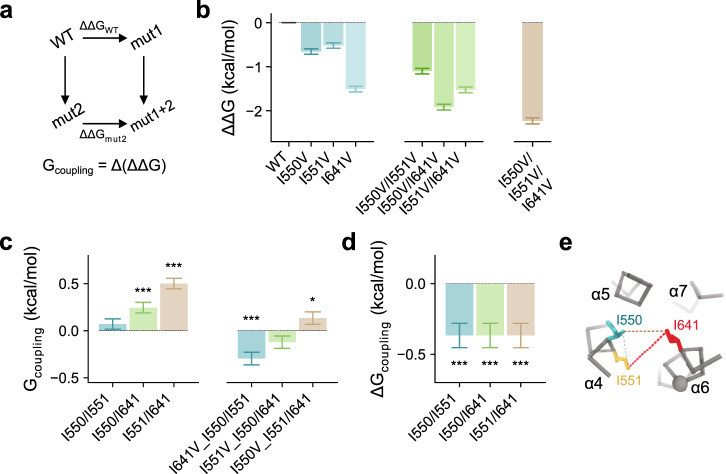
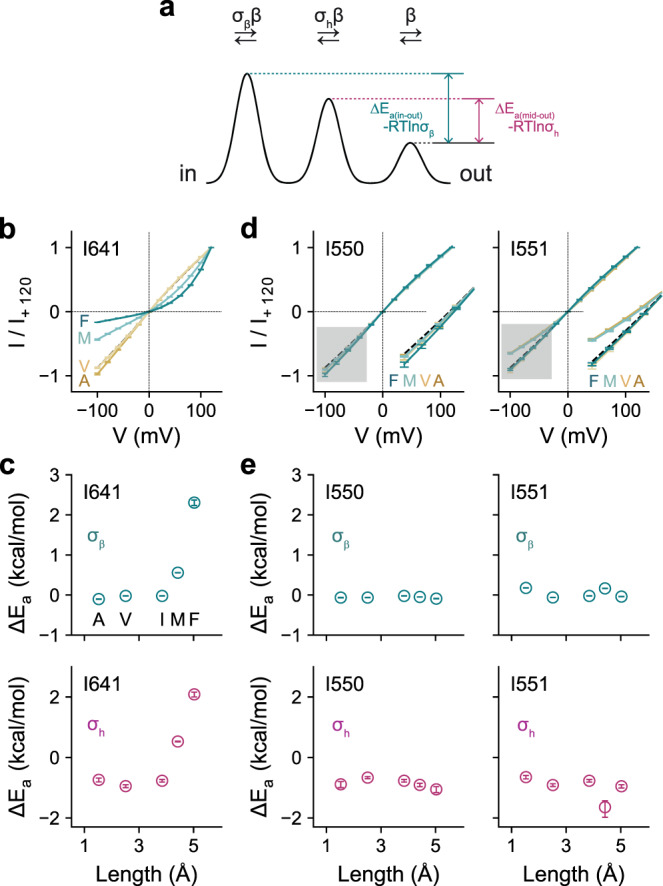
The binding of cytoplasmic Ca2+ to the anion-selective channel TMEM16A triggers a conformational change around its binding site that is coupled to the release of a gate at the constricted neck of an hourglass-shaped pore. By combining mutagenesis, electrophysiology, and cryo-electron microscopy, we identified three hydrophobic residues at the intracellular entrance of the neck as constituents of this gate. Mutation of each of these residues increases the potency of Ca2+ and results in pronounced basal activity. The structure of an activating mutant shows a conformational change of an α-helix that contributes to Ca2+ binding as a likely cause for the basal activity. Although not in physical contact, the three residues are functionally coupled to collectively contribute to the stabilization of the gate in the closed conformation of the pore, thus explaining the low open probability of the channel in the absence of Ca2+.
Conflict of interest statement
The authors declare no competing interests.
Figures


Similar articles
-
Mechanism of pore opening in the calcium-activated chloride channel TMEM16A.Nat Commun. 2021 Feb 4;12(1):786. doi: 10.1038/s41467-020-20788-8. Nat Commun. 2021. PMID: 33542228 Free PMC article.
-
Cryo-EM structures of the TMEM16A calcium-activated chloride channel.Nature. 2017 Dec 21;552(7685):426-429. doi: 10.1038/nature25024. Epub 2017 Dec 13. Nature. 2017. PMID: 29236684 Free PMC article.
-
Voltage-Dependent Protonation of the Calcium Pocket Enable Activation of the Calcium-Activated Chloride Channel Anoctamin-1 (TMEM16A).Sci Rep. 2020 Apr 20;10(1):6644. doi: 10.1038/s41598-020-62860-9. Sci Rep. 2020. PMID: 32313203 Free PMC article.
-
Insights into the function and regulation of the calcium-activated chloride channel TMEM16A.Cell Calcium. 2024 Jul;121:102891. doi: 10.1016/j.ceca.2024.102891. Epub 2024 May 8. Cell Calcium. 2024. PMID: 38772195 Review.
-
Wasted TMEM16A channels are rescued by phosphatidylinositol 4,5-bisphosphate.Cell Calcium. 2019 Dec;84:102103. doi: 10.1016/j.ceca.2019.102103. Epub 2019 Oct 18. Cell Calcium. 2019. PMID: 31683182 Free PMC article. Review.
Cited by
-
Distinct modulation of calcium-activated chloride channel TMEM16A by drug-binding sites.Proc Natl Acad Sci U S A. 2024 Dec 17;121(51):e2314011121. doi: 10.1073/pnas.2314011121. Epub 2024 Dec 10. Proc Natl Acad Sci U S A. 2024. PMID: 39656212 Free PMC article.
-
TMEM16 and OSCA/TMEM63 proteins share a conserved potential to permeate ions and phospholipids.bioRxiv [Preprint]. 2024 Sep 3:2024.02.04.578431. doi: 10.1101/2024.02.04.578431. bioRxiv. 2024. Update in: Elife. 2024 Nov 04;13:RP96957. doi: 10.7554/eLife.96957. PMID: 38370744 Free PMC article. Updated. Preprint.
-
Gating and Regulatory Mechanisms of TMEM16 Ion Channels and Scramblases.Front Physiol. 2021 Nov 19;12:787773. doi: 10.3389/fphys.2021.787773. eCollection 2021. Front Physiol. 2021. PMID: 34867487 Free PMC article. Review.
-
Computational Investigation of Mechanisms for pH Modulation of Human Chloride Channels.Molecules. 2023 Jul 30;28(15):5753. doi: 10.3390/molecules28155753. Molecules. 2023. PMID: 37570721 Free PMC article.
-
Mechanistic basis of ligand efficacy in the calcium-activated chloride channel TMEM16A.EMBO J. 2023 Dec 11;42(24):e115030. doi: 10.15252/embj.2023115030. Epub 2023 Nov 20. EMBO J. 2023. PMID: 37984335 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
