

Membrane-destabilizing ionizable phospholipids for organ-selective mRNA delivery and CRISPR-Cas gene editing
- PMID: 33542471
- PMCID: PMC8188687
- DOI: 10.1038/s41563-020-00886-0
Membrane-destabilizing ionizable phospholipids for organ-selective mRNA delivery and CRISPR-Cas gene editing
Abstract
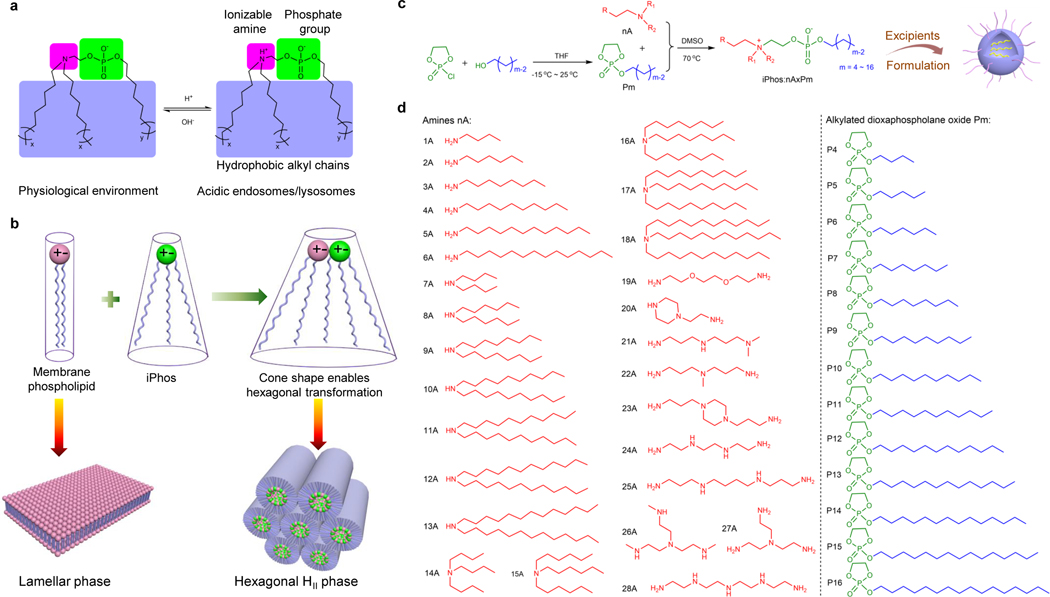
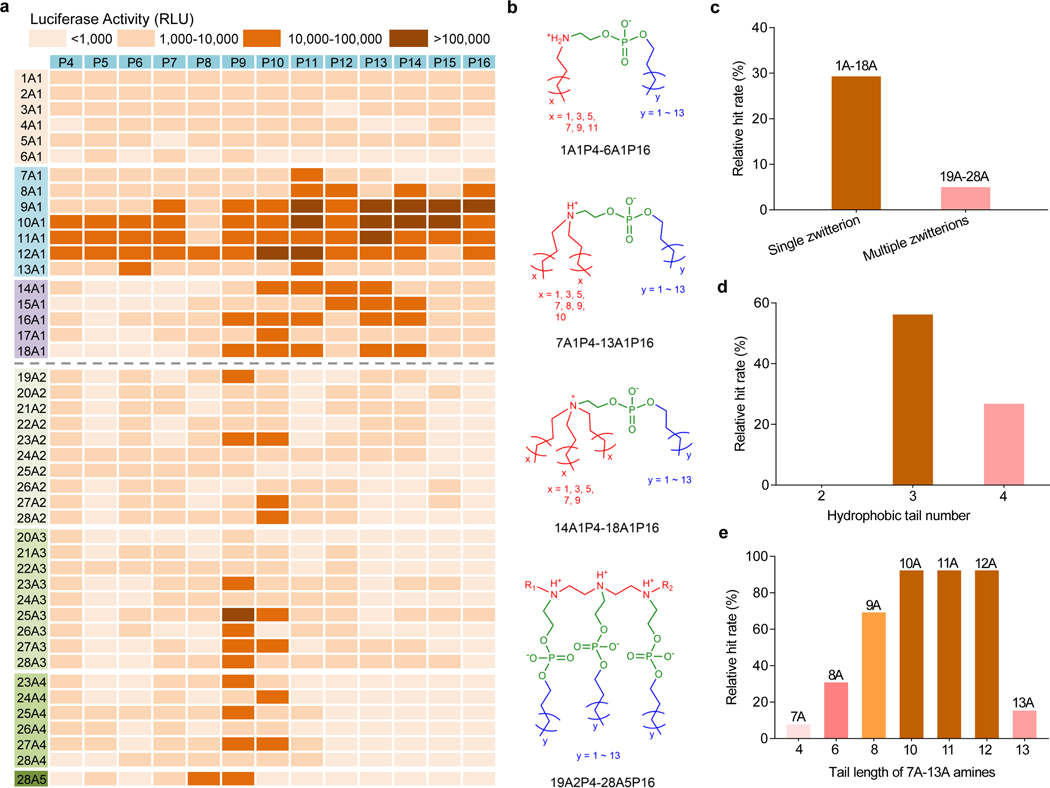
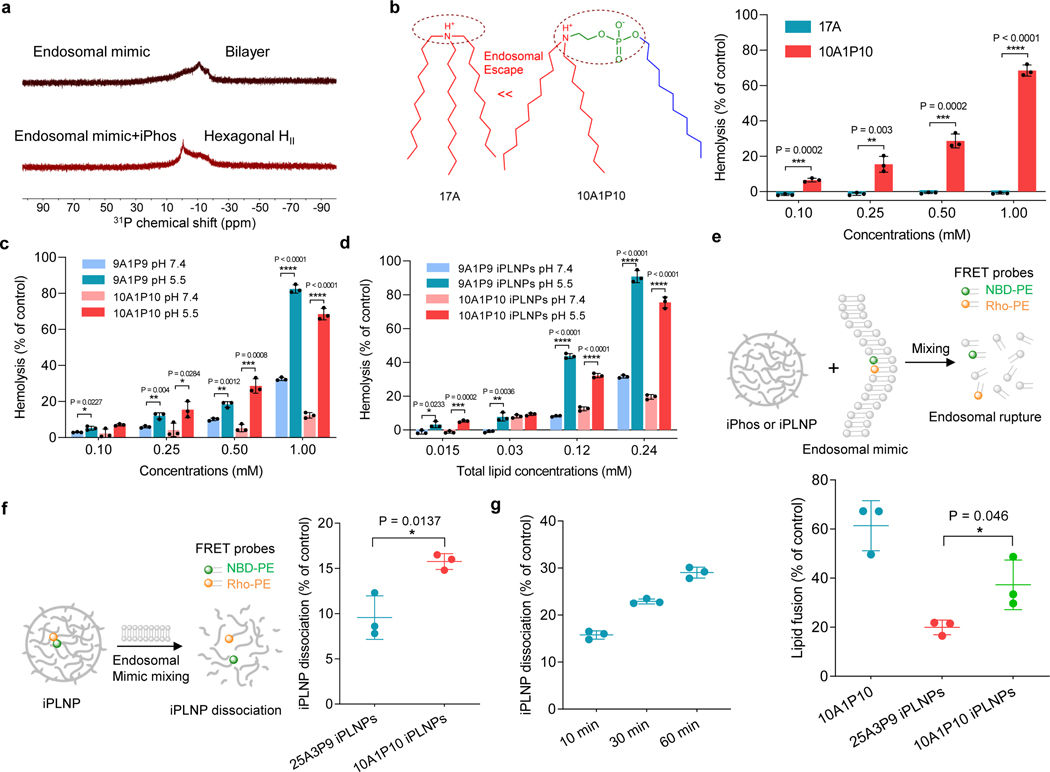
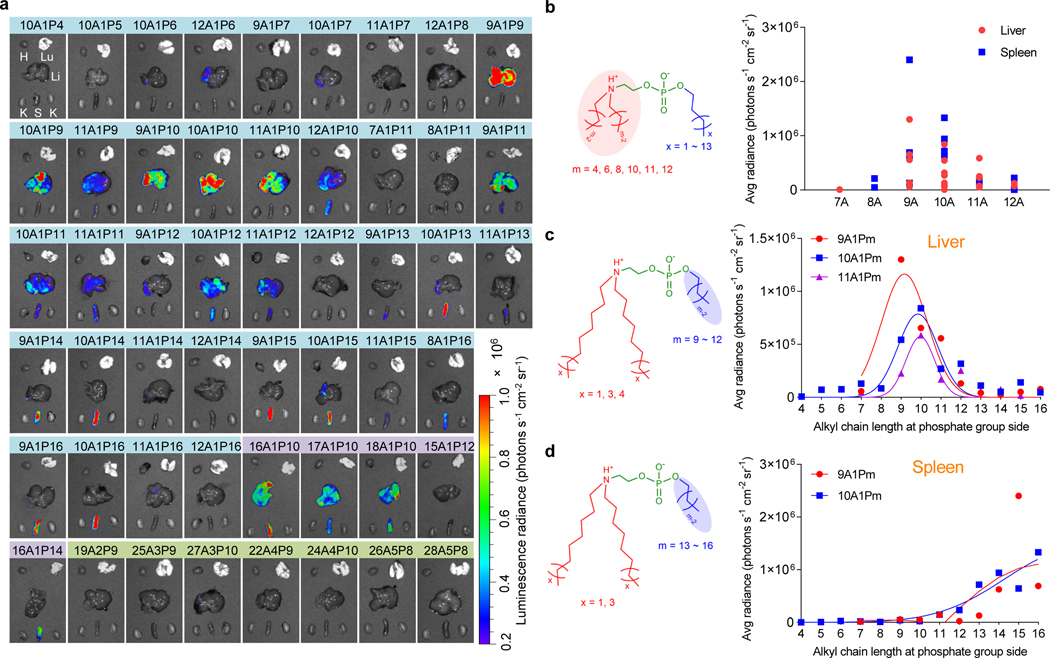
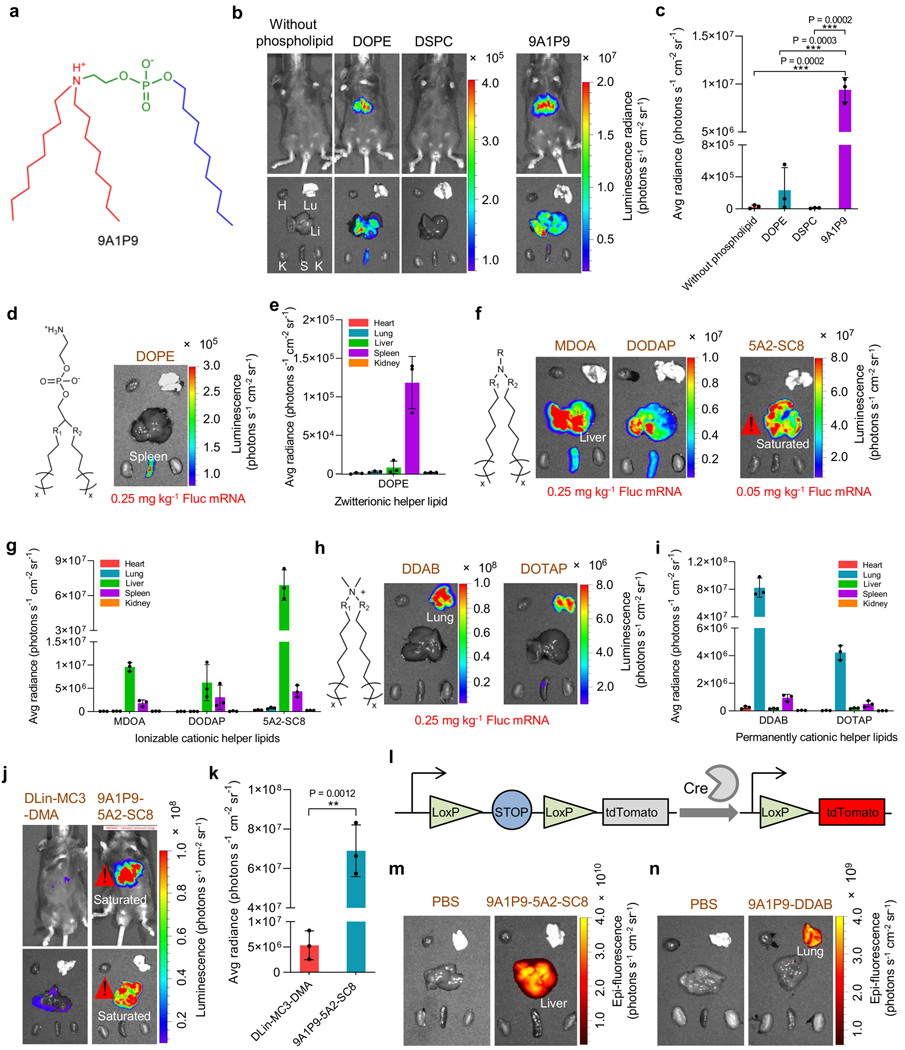
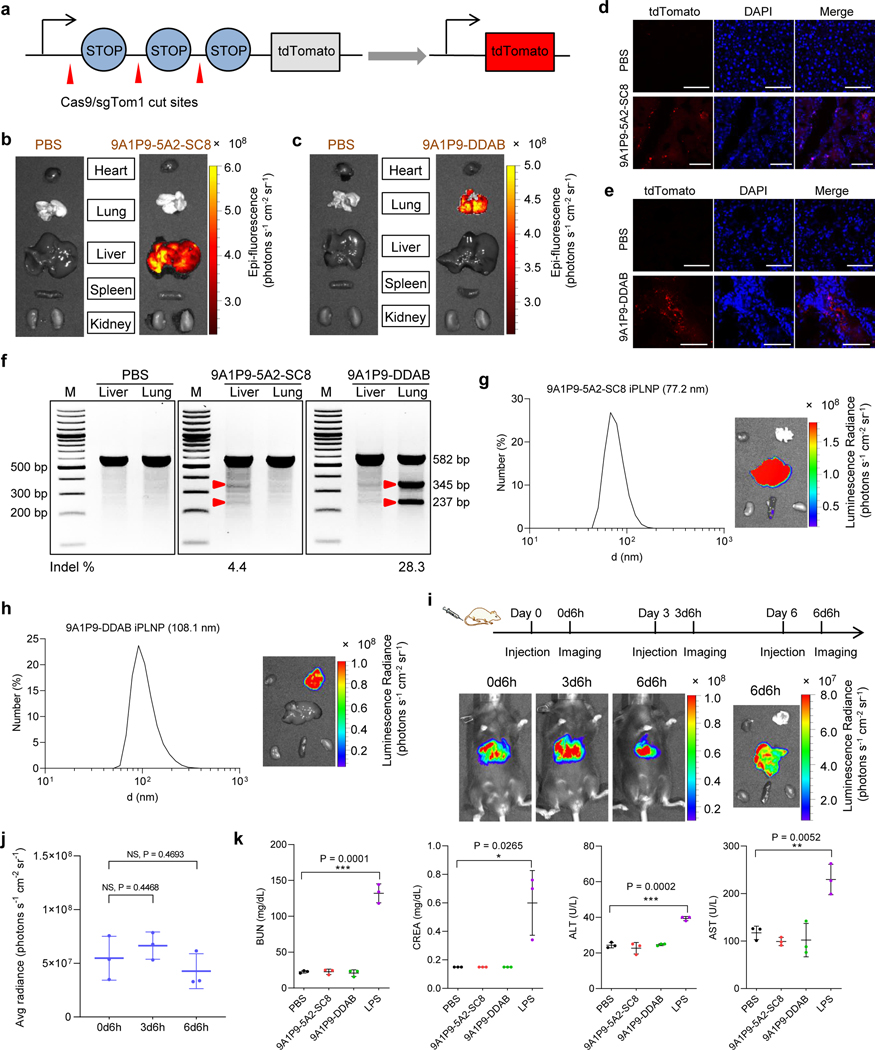
Endosomal escape remains a fundamental barrier hindering the advancement of nucleic acid therapeutics. Taking inspiration from natural phospholipids that comprise biological membranes, we report the combinatorial synthesis of multi-tailed ionizable phospholipids (iPhos) capable of delivering messenger RNA or mRNA/single-guide RNA for gene editing in vivo. Optimized iPhos lipids are composed of one pH-switchable zwitterion and three hydrophobic tails, which adopt a cone shape in endosomal acidic environments to facilitate membrane hexagonal transformation and subsequent cargo release from endosomes. Structure-activity relationships reveal that iPhos chemical structure can control in vivo efficacy and organ selectivity. iPhos lipids synergistically function with various helper lipids to formulate multi-component lipid nanoparticles (called iPLNPs) for selective organ targeting. Zwitterionic, ionizable cationic and permanently cationic helper lipids enable tissue-selective mRNA delivery and CRISPR-Cas9 gene editing in spleen, liver and lungs (respectively) following intravenous administration. This rational design of functional phospholipids demonstrates substantial value for gene editing research and therapeutic applications.
Figures
Comment in
-
Overcoming delivery barriers with LNPs.Nat Mater. 2021 May;20(5):575-577. doi: 10.1038/s41563-021-00988-3. Nat Mater. 2021. PMID: 33911220 No abstract available.
References
-
- Wang HX et al. CRISPR/Cas9-based genome editing for disease modeling and therapy: Challenges and opportunities for nonviral delivery. Chem. Rev 117, 9874–9906 (2017). - PubMed
-
- Hajj KA & Whitehead KA Tools for translation: Non-viral materials for therapeutic mRNA delivery. Nat. Rev. Mater 2, 17056 (2017).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
