

CryoDRGN: reconstruction of heterogeneous cryo-EM structures using neural networks
- PMID: 33542510
- PMCID: PMC8183613
- DOI: 10.1038/s41592-020-01049-4
CryoDRGN: reconstruction of heterogeneous cryo-EM structures using neural networks
Abstract
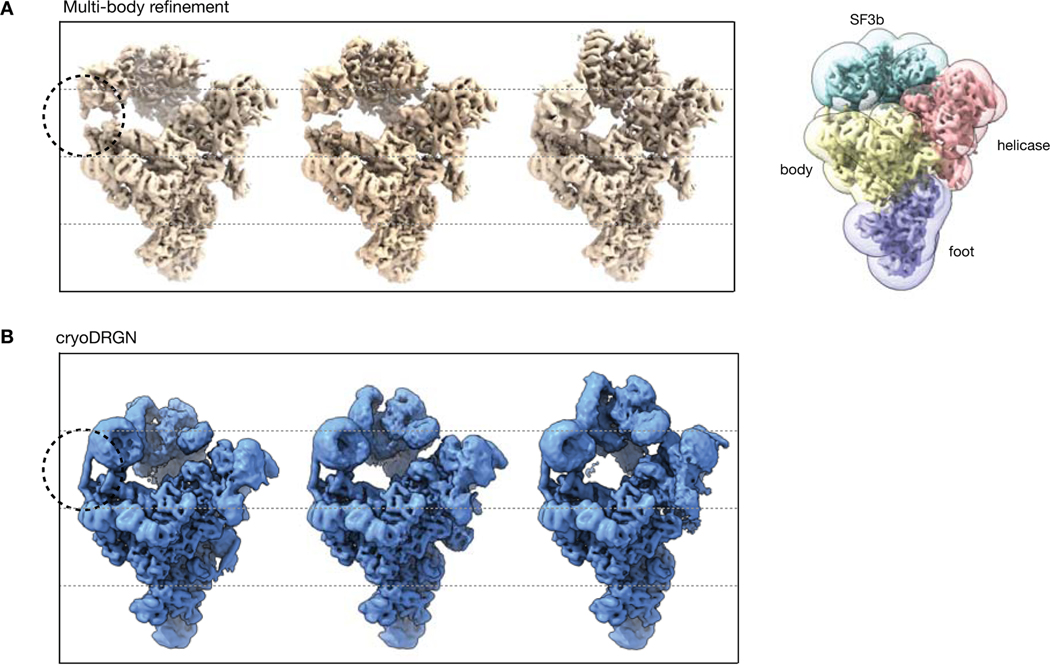
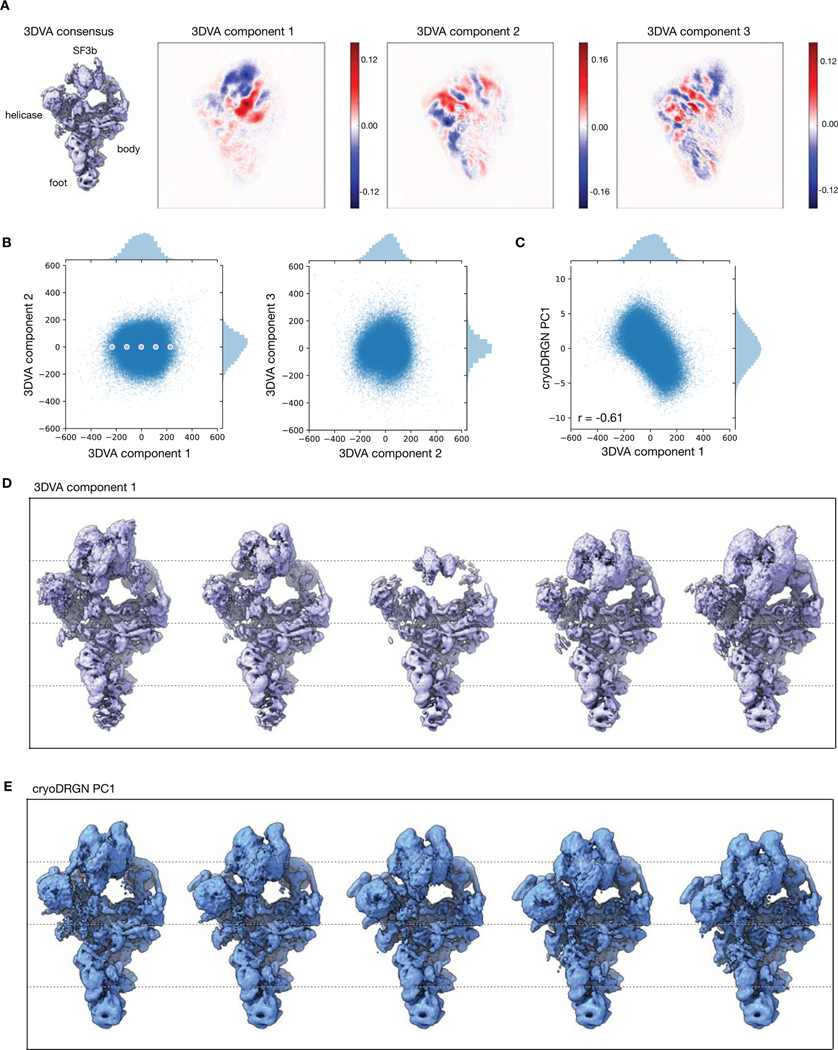
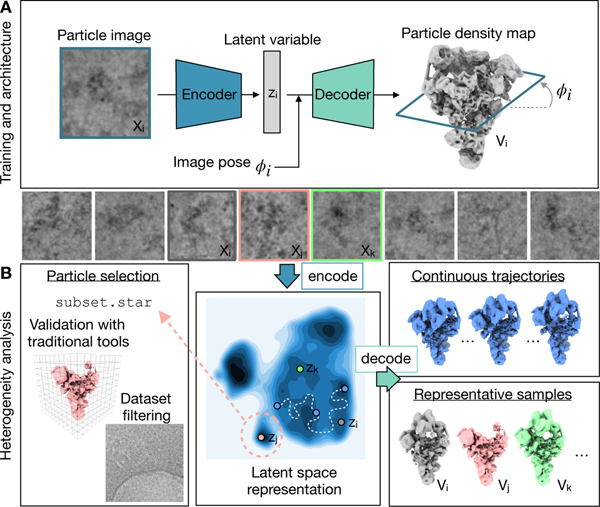
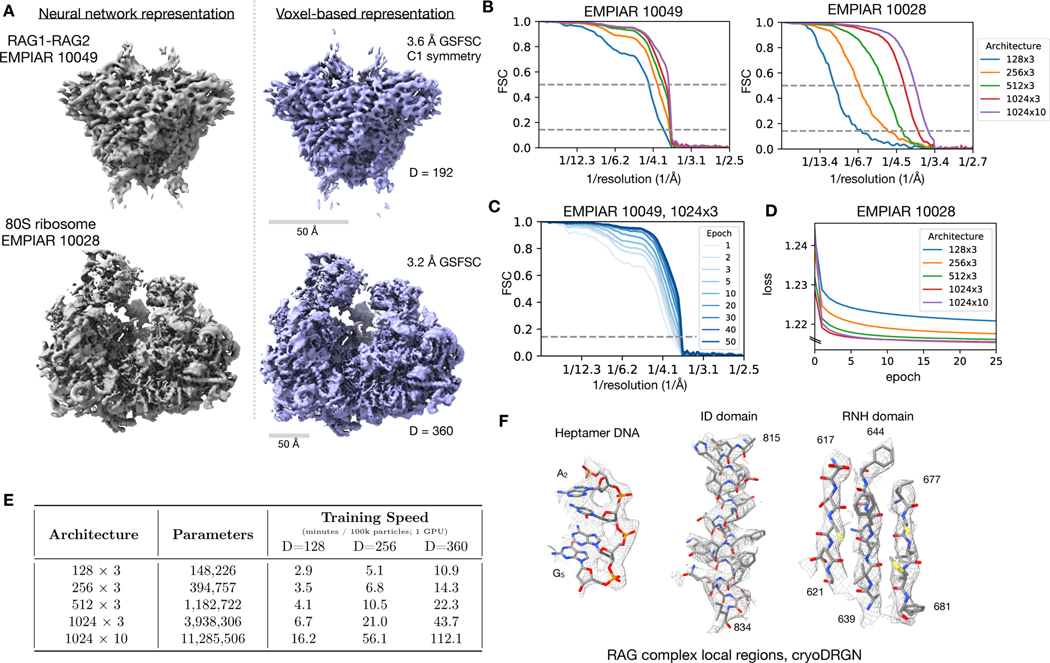
Cryo-electron microscopy (cryo-EM) single-particle analysis has proven powerful in determining the structures of rigid macromolecules. However, many imaged protein complexes exhibit conformational and compositional heterogeneity that poses a major challenge to existing three-dimensional reconstruction methods. Here, we present cryoDRGN, an algorithm that leverages the representation power of deep neural networks to directly reconstruct continuous distributions of 3D density maps and map per-particle heterogeneity of single-particle cryo-EM datasets. Using cryoDRGN, we uncovered residual heterogeneity in high-resolution datasets of the 80S ribosome and the RAG complex, revealed a new structural state of the assembling 50S ribosome, and visualized large-scale continuous motions of a spliceosome complex. CryoDRGN contains interactive tools to visualize a dataset's distribution of per-particle variability, generate density maps for exploratory analysis, extract particle subsets for use with other tools and generate trajectories to visualize molecular motions. CryoDRGN is open-source software freely available at http://cryodrgn.csail.mit.edu .
Conflict of interest statement
Ethics Declaration
The authors declare no competing financial interests.
Figures
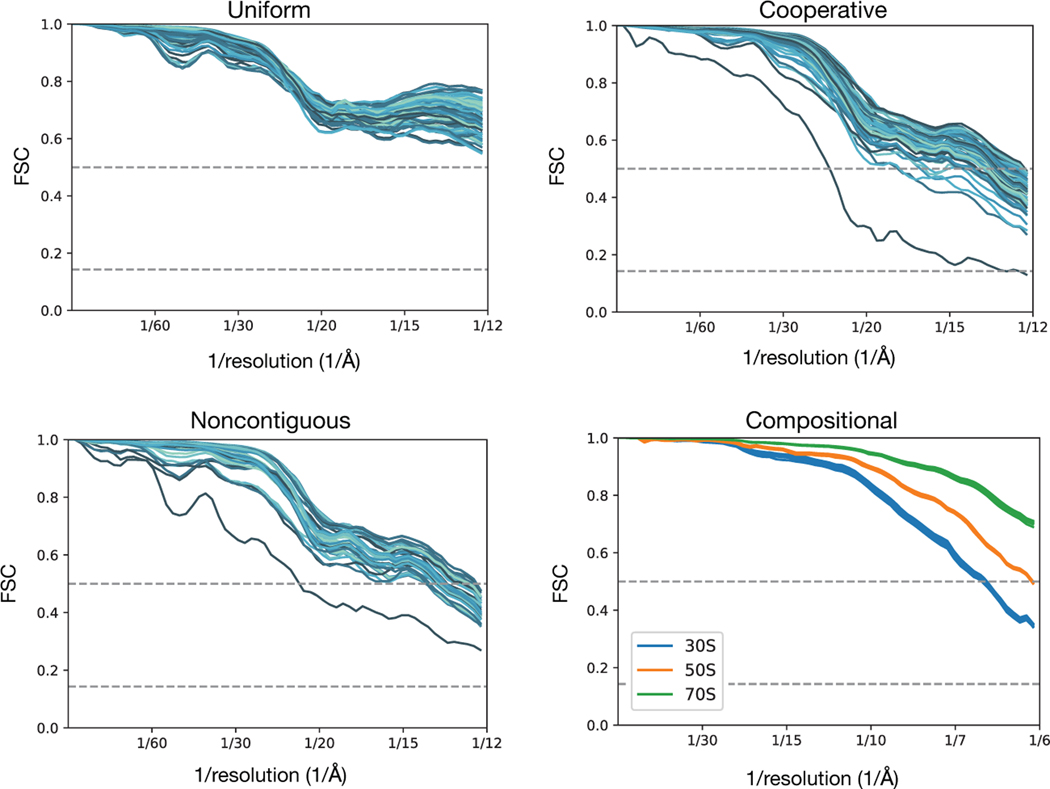
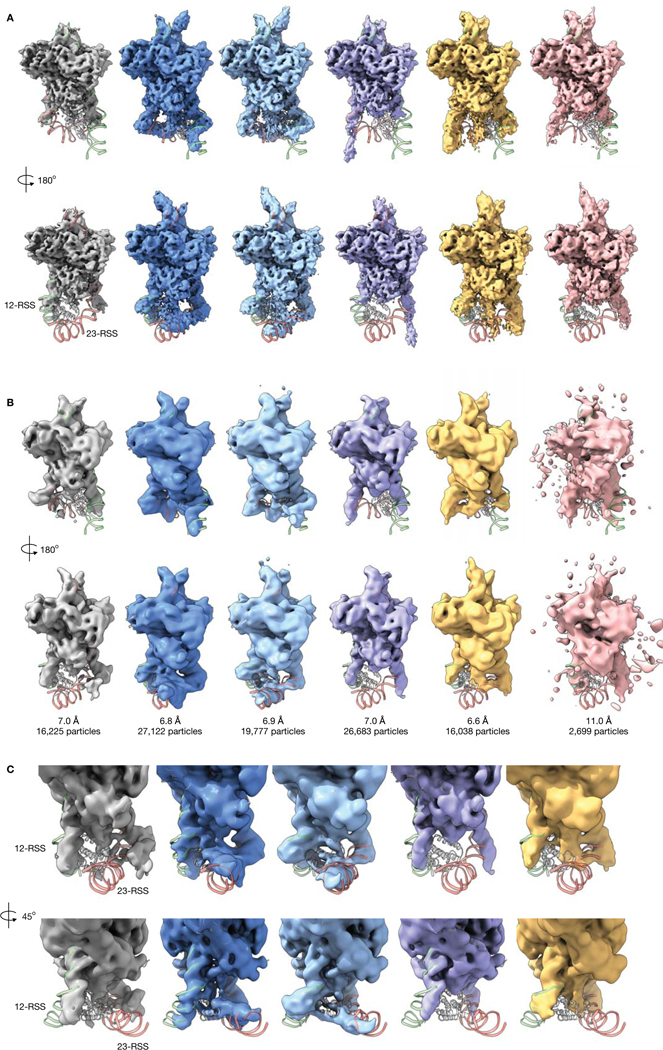
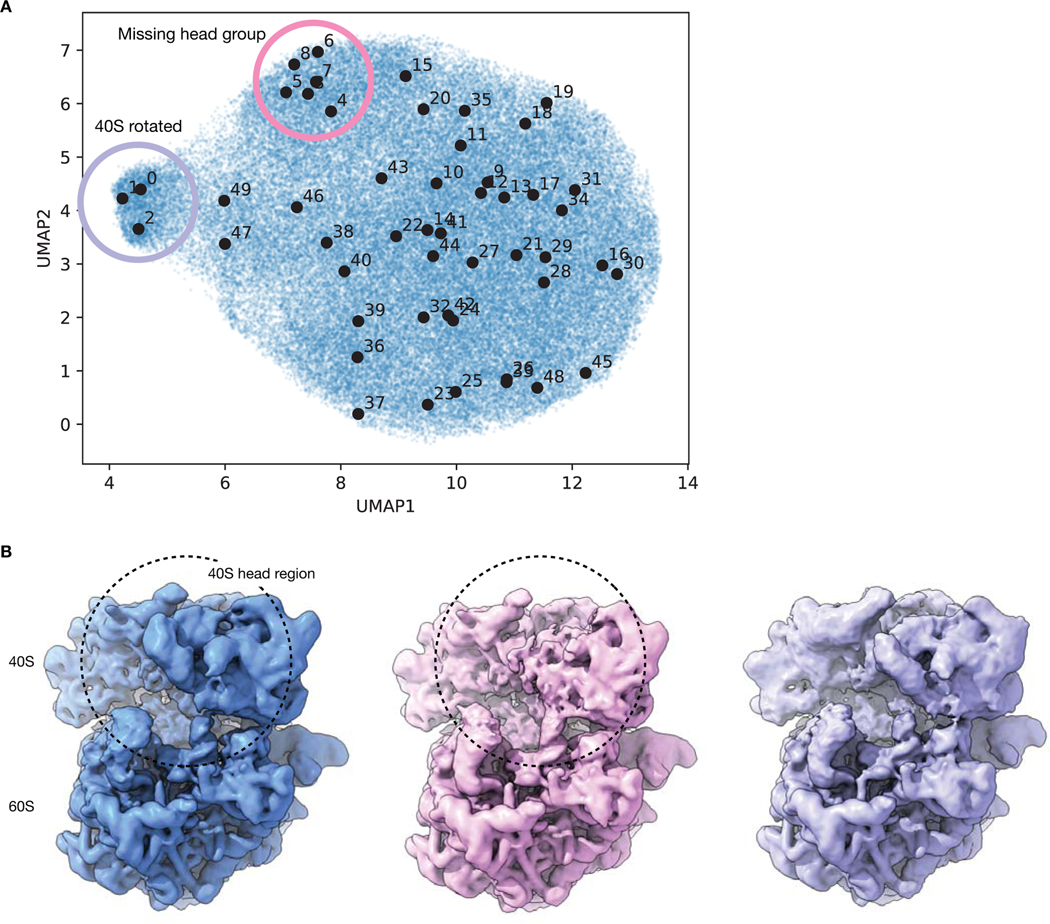
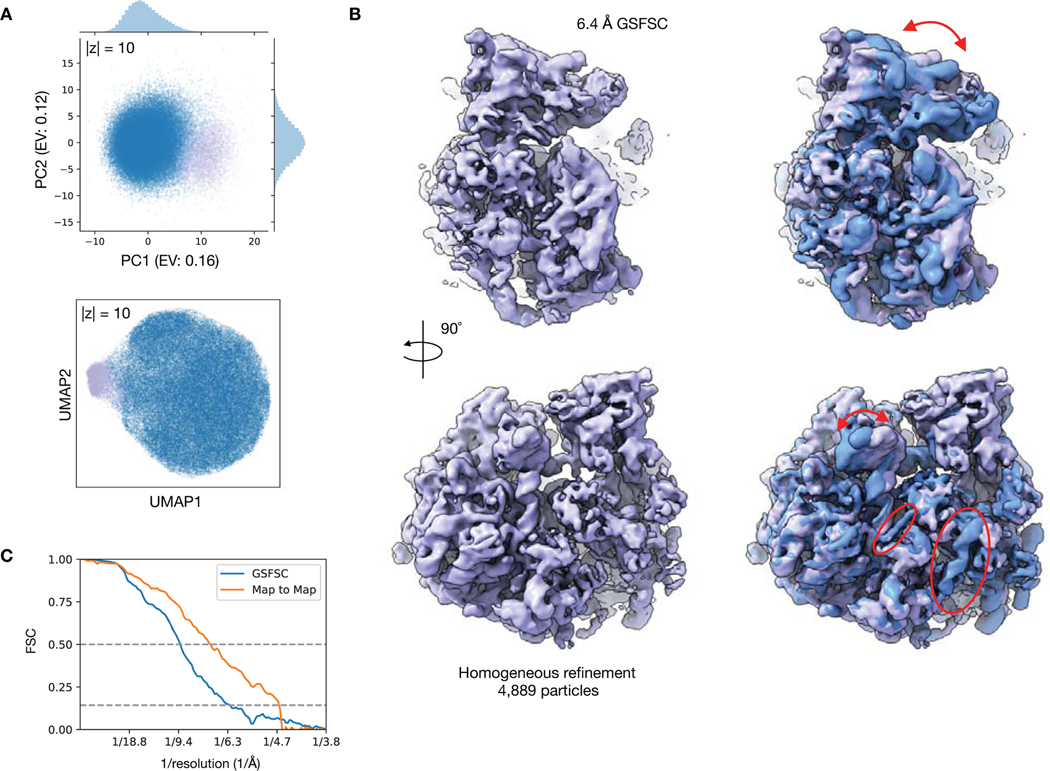
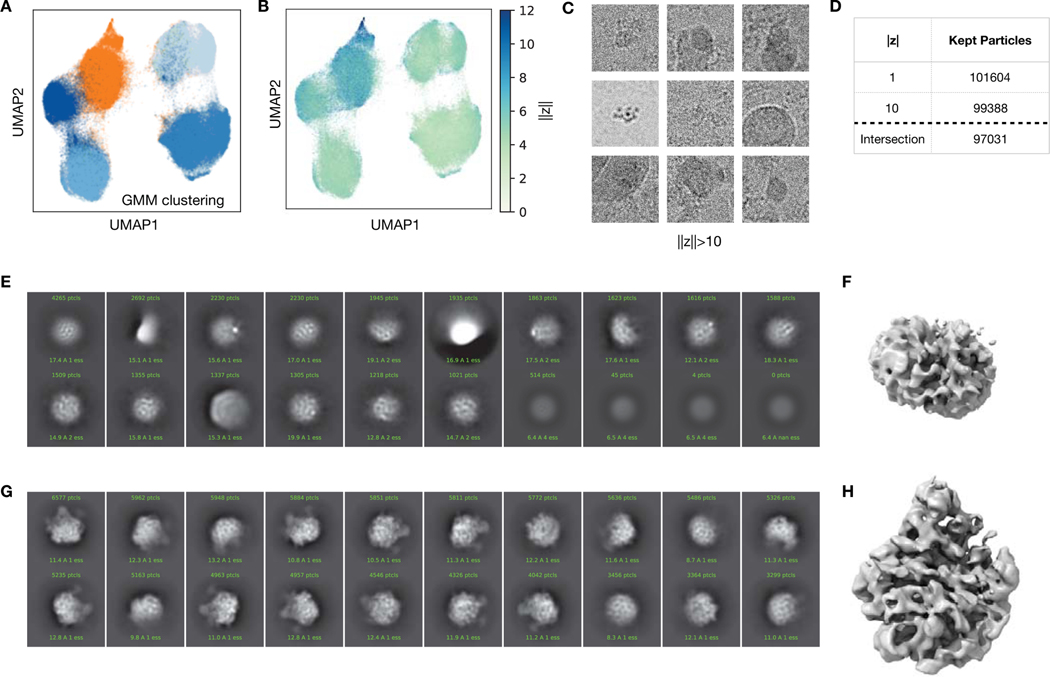
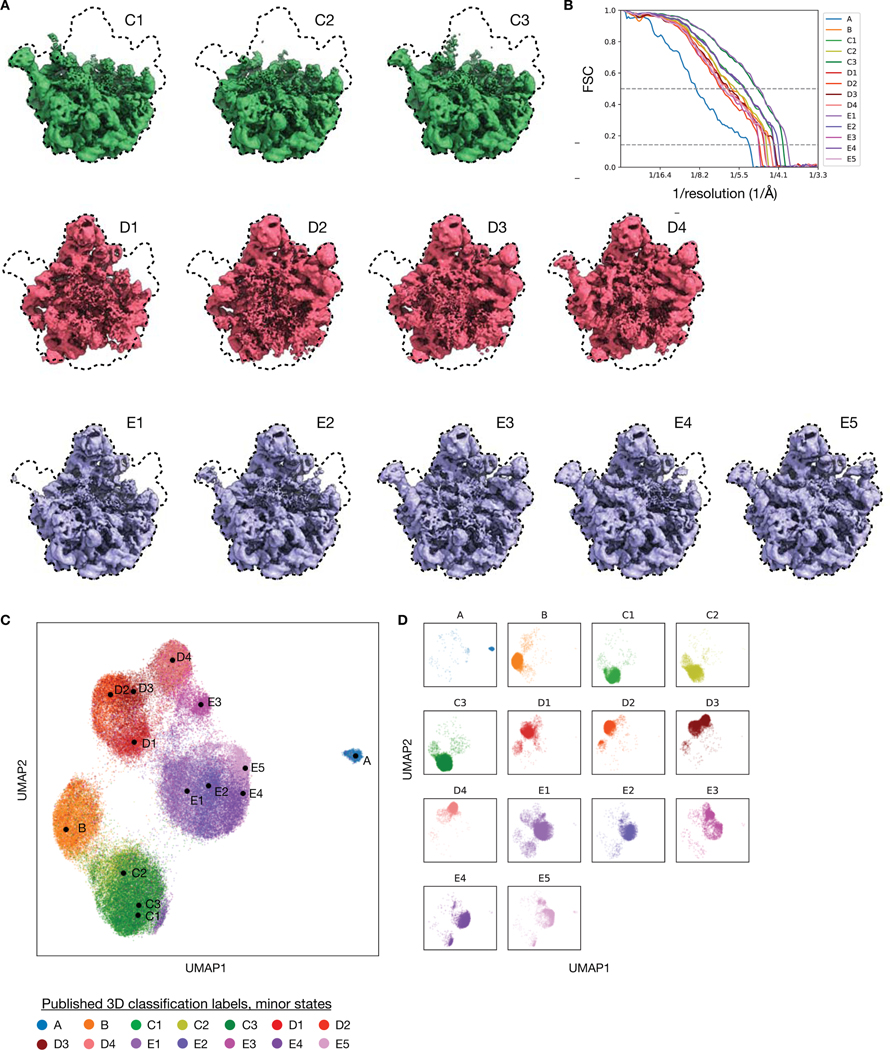
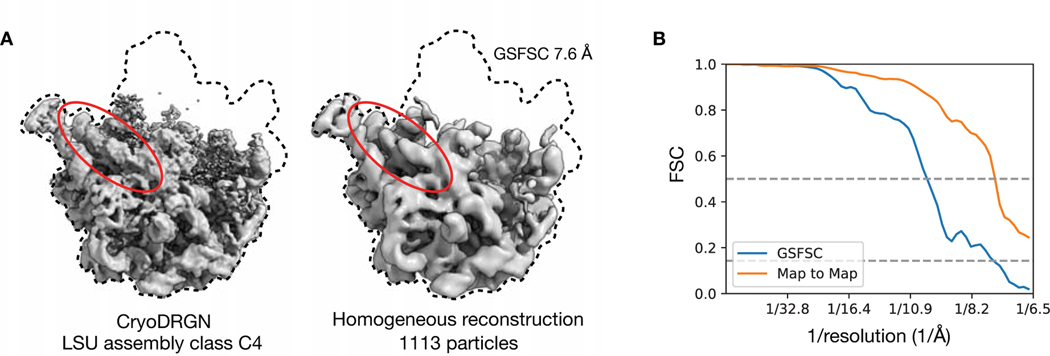
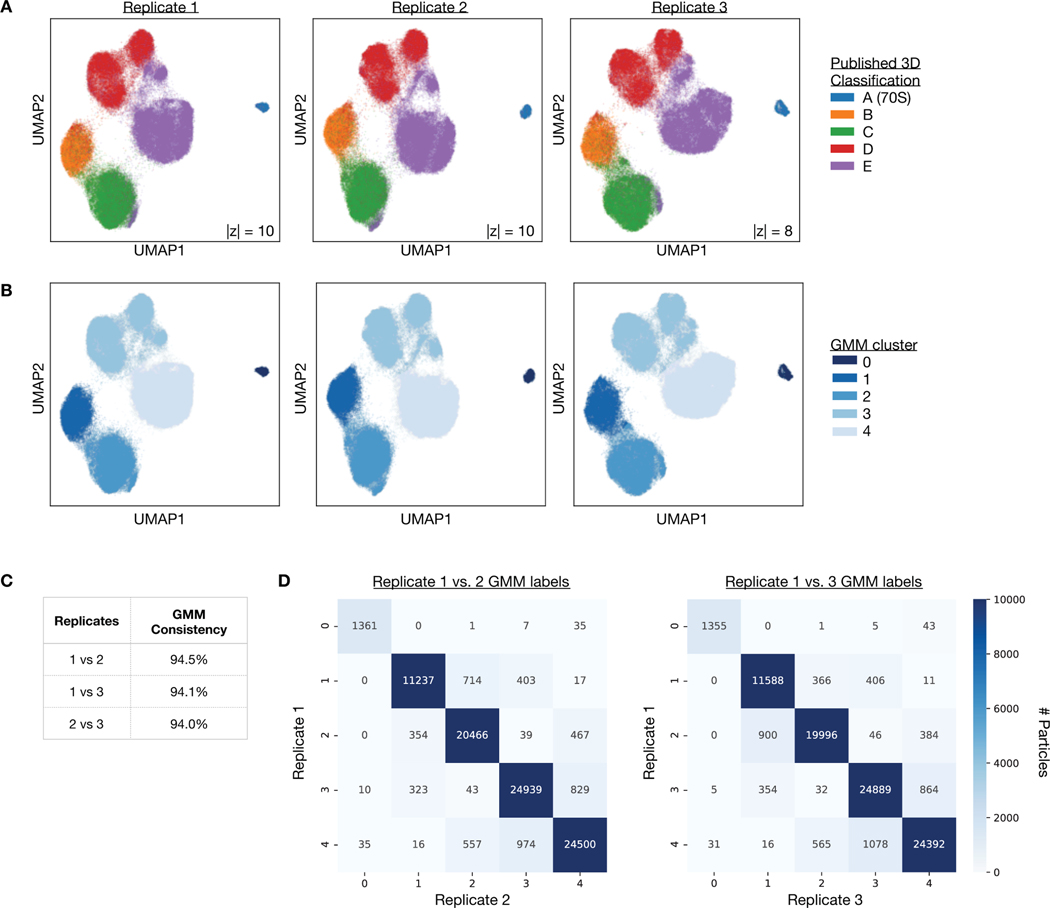
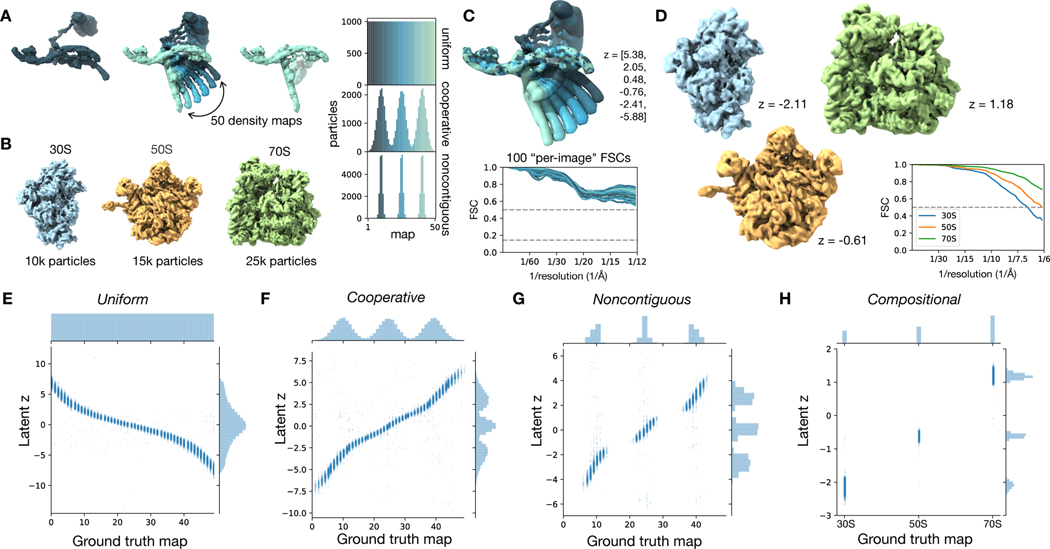
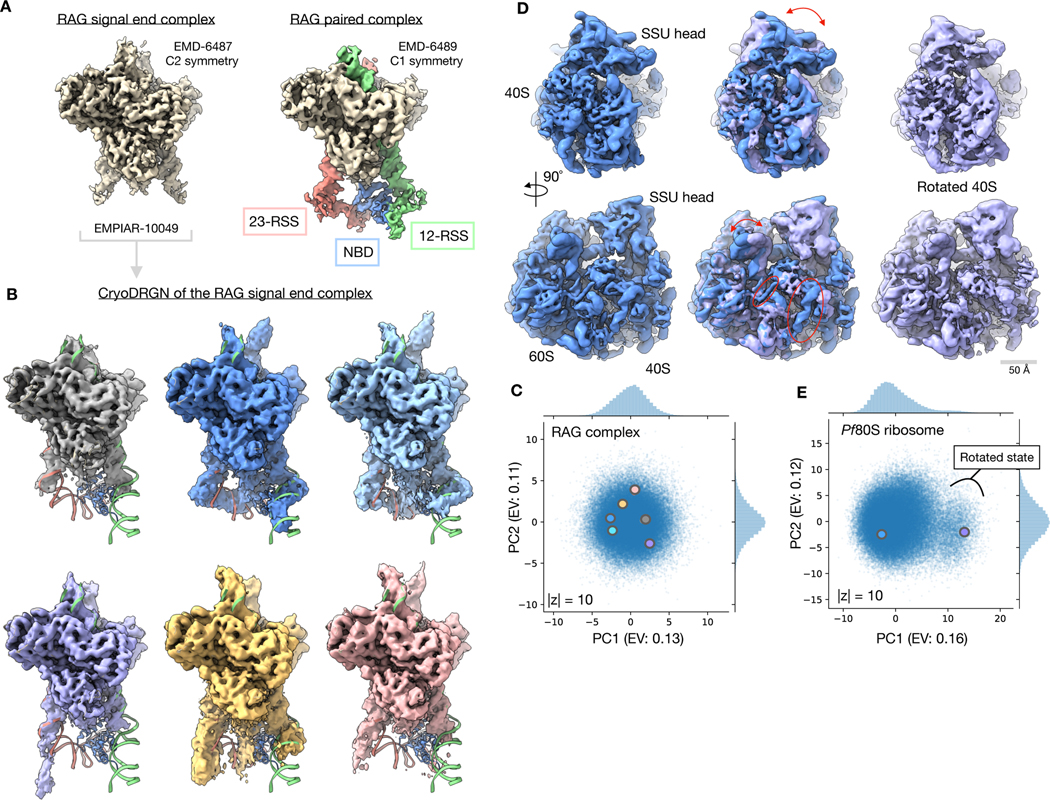
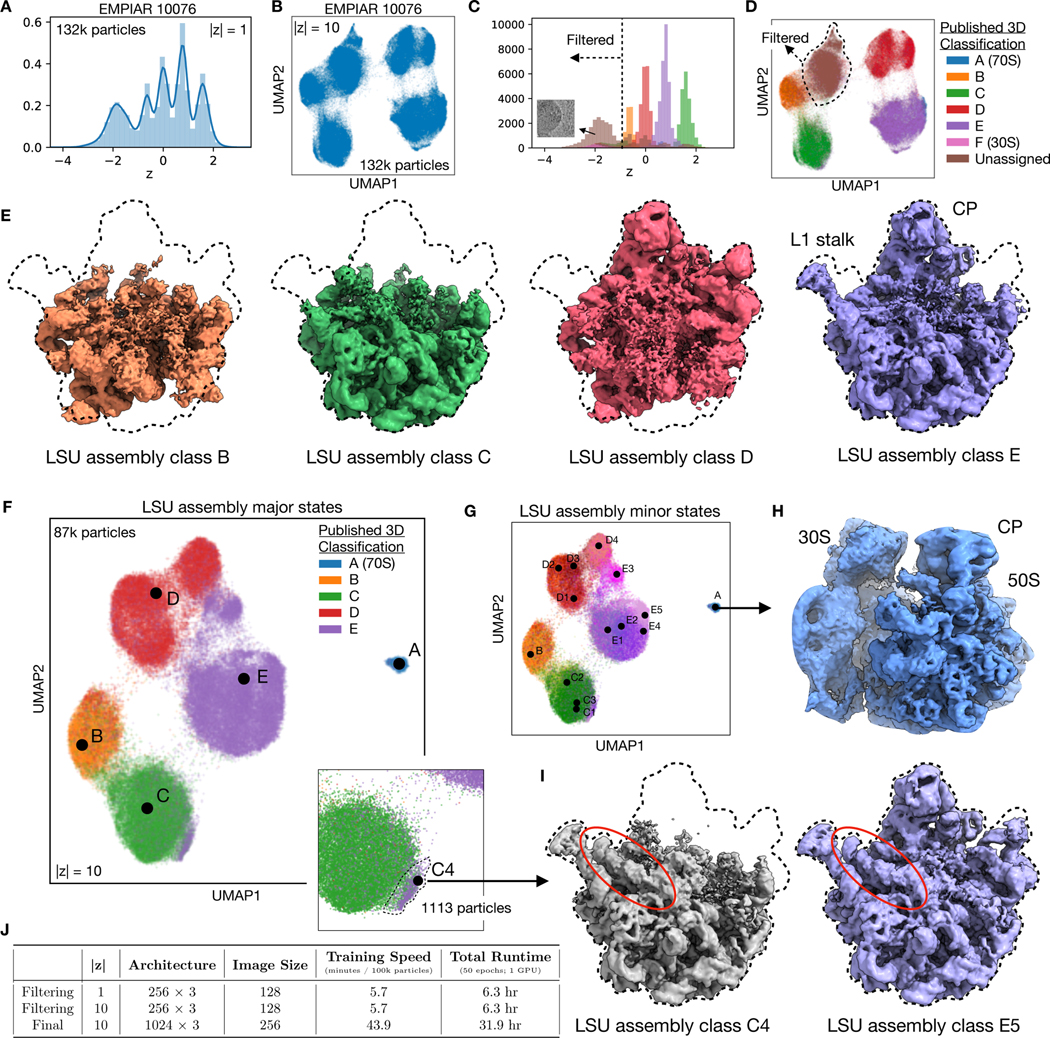
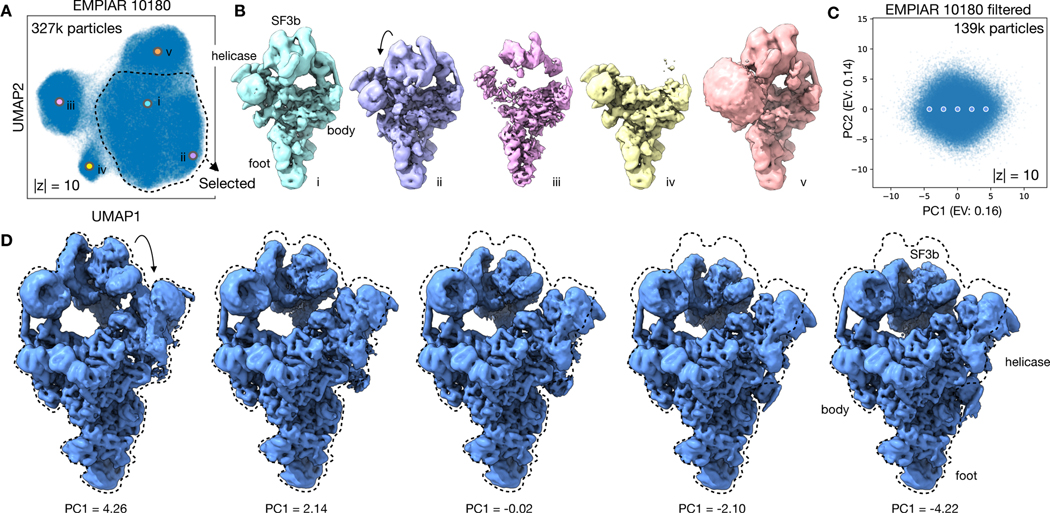
Comment in
-
Neural networks learn the motions of molecular machines.Nat Methods. 2021 Aug;18(8):869-871. doi: 10.1038/s41592-021-01235-y. Nat Methods. 2021. PMID: 34326542 No abstract available.
References
-
- Suloway C. et al. Automated molecular microscopy: The new Leginon system. J. Struct. Biol 151, 41–60 (2005). - PubMed
Methods-only References
-
- Zhong ED, Bepler T, Davis JH & Berger B. Reconstructing continuous distributions of 3 D protein structure from cryo-EM images in International Conference of Learning Representations, ICLR (2020).
-
- Bepler T, Zhong E, Kelley K, Brignole E. & Berger B. Explicitly disentangling image content from translation and rotation with spatial-VAE. in Advances in Neural Information Processing Systems (2019).
-
- Vaswani A. et al. Attention is all you need. in Advances in Neural Information Processing Systems (2017).
-
- Rezende DJ, Mohamed S. & Wierstra D. Stochastic backpropagation and approximate inference in deep generative models. in International Conference on Machine Learning, ICML (2014).
-
- The PyMOL Molecular Graphics System, Version 2.3, Schrodinger, LLC.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
