

C9orf72 regulates energy homeostasis by stabilizing mitochondrial complex I assembly
- PMID: 33545050
- PMCID: PMC8579819
- DOI: 10.1016/j.cmet.2021.01.005
C9orf72 regulates energy homeostasis by stabilizing mitochondrial complex I assembly
Abstract
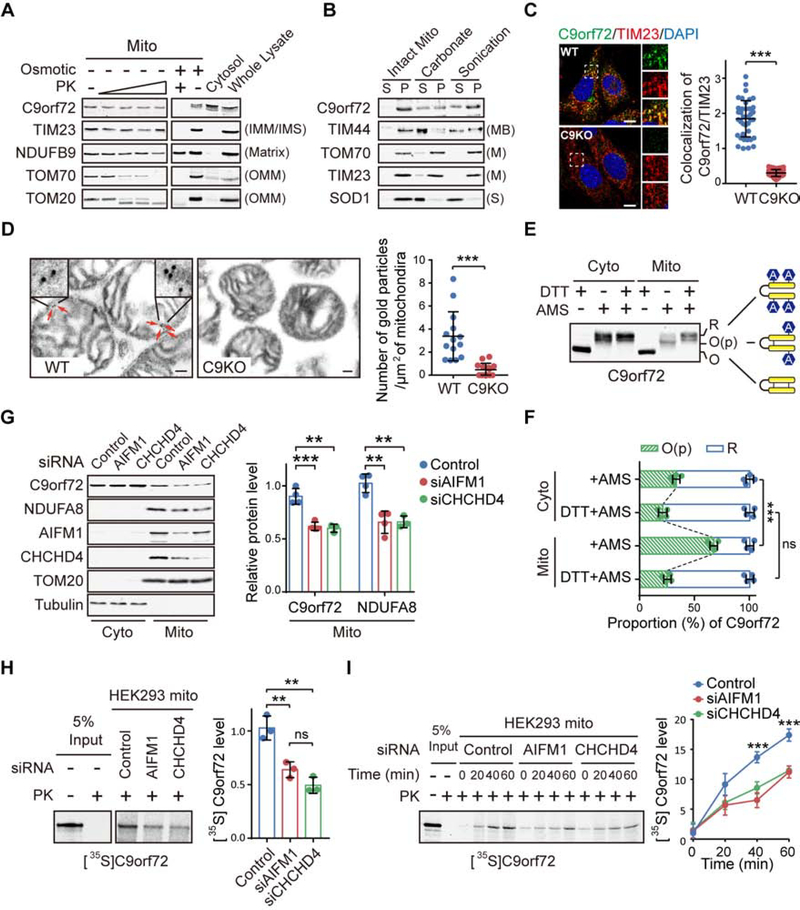
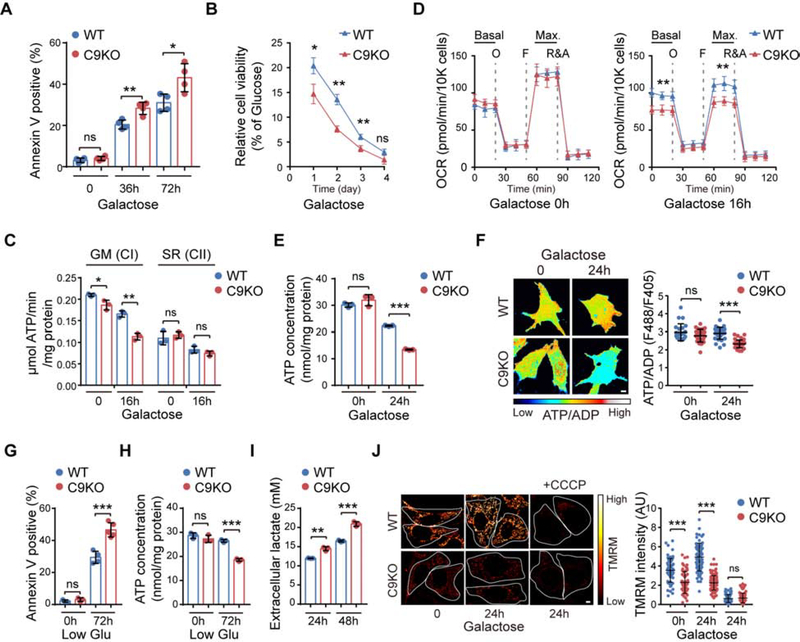
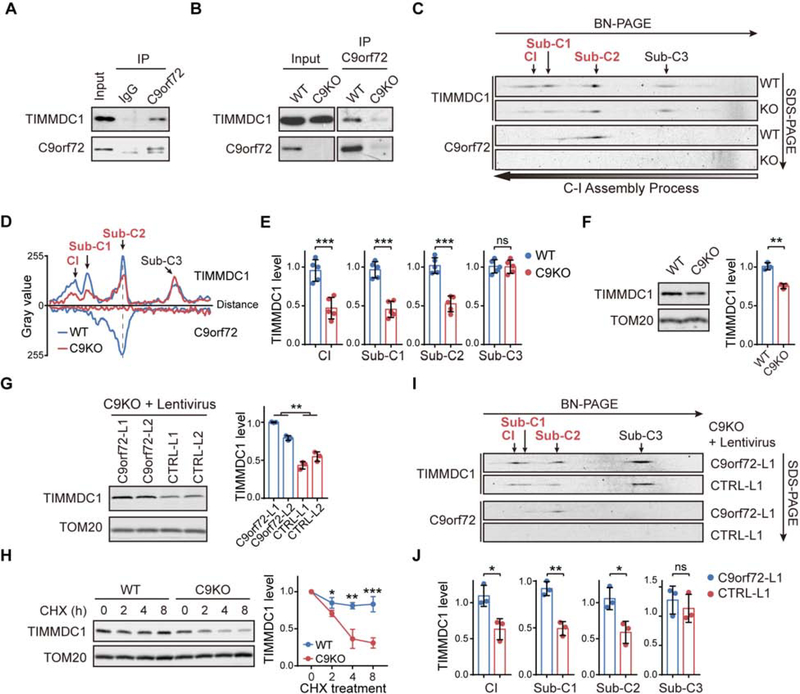
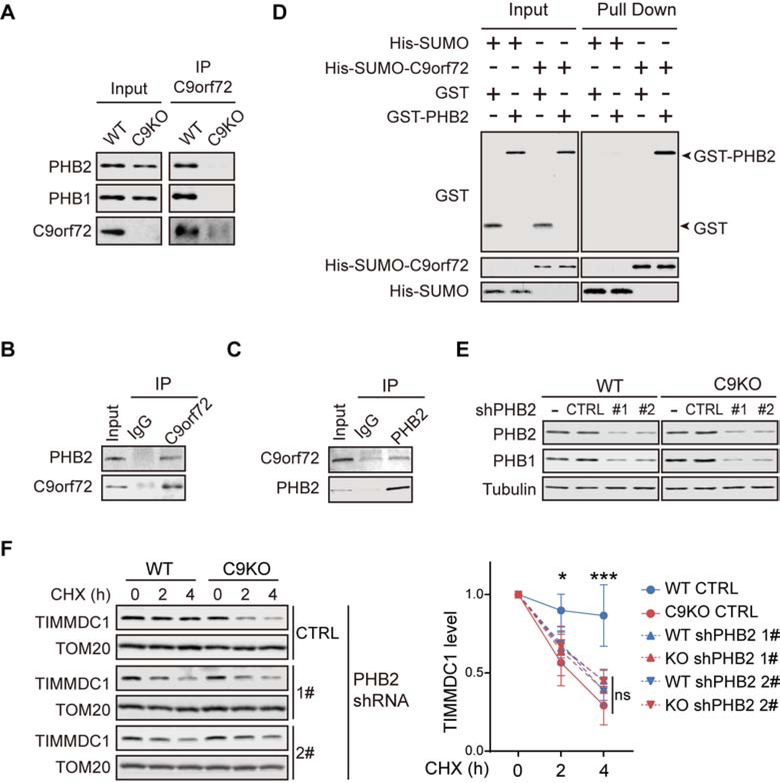
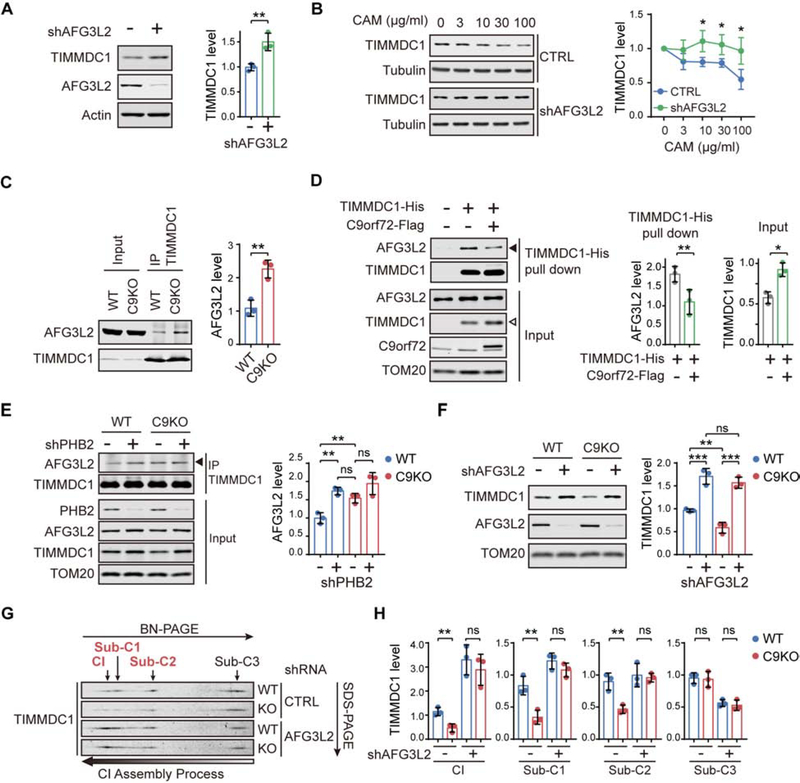
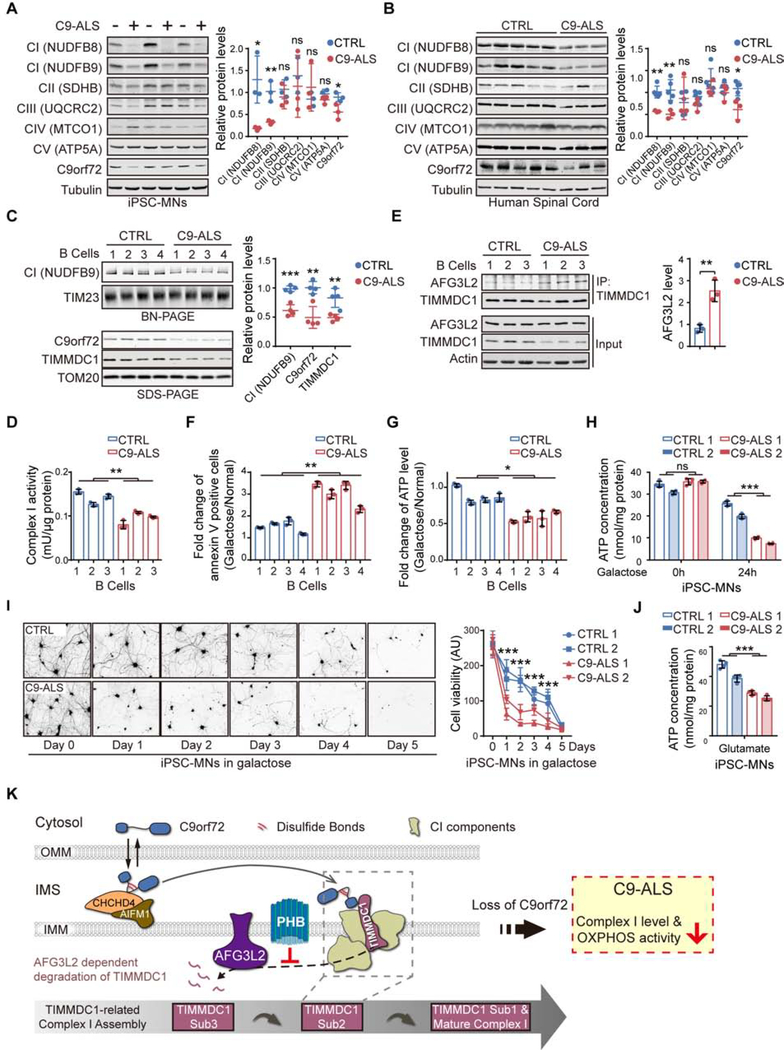
The haploinsufficiency of C9orf72 is implicated in the most common forms of amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD), but the full spectrum of C9orf72 functions remains to be established. Here, we report that C9orf72 is a mitochondrial inner-membrane-associated protein regulating cellular energy homeostasis via its critical role in the control of oxidative phosphorylation (OXPHOS). The translocation of C9orf72 from the cytosol to the inter-membrane space is mediated by the redox-sensitive AIFM1/CHCHD4 pathway. In mitochondria, C9orf72 specifically stabilizes translocase of inner mitochondrial membrane domain containing 1 (TIMMDC1), a crucial factor for the assembly of OXPHOS complex I. C9orf72 directly recruits the prohibitin complex to inhibit the m-AAA protease-dependent degradation of TIMMDC1. The mitochondrial complex I function is impaired in C9orf72-linked ALS/FTD patient-derived neurons. These results reveal a previously unknown function of C9orf72 in mitochondria and suggest that defective energy metabolism may underlie the pathogenesis of relevant diseases.
Keywords: ALS; C9orf72; FTD; OXPHOS; TIMMDC1; complex I; mitochondrial import; mitochondrion; neurodegeneration; oxidative phosphorylation.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures

References
-
- Acin-Perez R, Fernandez-Silva P, Peleato ML, Perez-Martos A, and Enriquez JA (2008). Respiratory active mitochondrial supercomplexes. Molecular cell 32, 529–539. - PubMed
-
- Dafinca R, Scaber J, Ababneh N, Lalic T, Weir G, Christian H, Vowles J, Douglas AG, Fletcher-Jones A, Browne C, et al. (2016). C9orf72 Hexanucleotide Expansions Are Associated with Altered Endoplasmic Reticulum Calcium Homeostasis and Stress Granule Formation in Induced Pluripotent Stem Cell-Derived Neurons from Patients with Amyotrophic Lateral Sclerosis and Frontotemporal Dementia. Stem Cells 34, 2063–2078. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
