

Functional analysis of sense organ specification in the Tribolium castaneum larva reveals divergent mechanisms in insects
- PMID: 33546687
- PMCID: PMC7866635
- DOI: 10.1186/s12915-021-00948-y
Functional analysis of sense organ specification in the Tribolium castaneum larva reveals divergent mechanisms in insects
Abstract
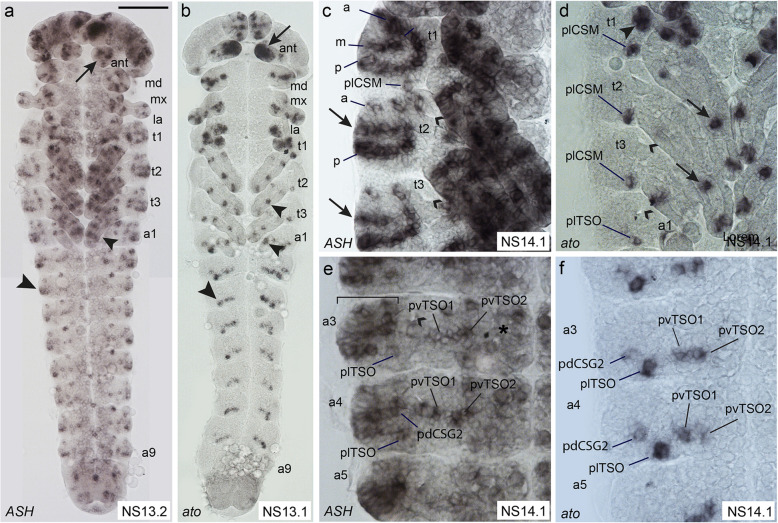
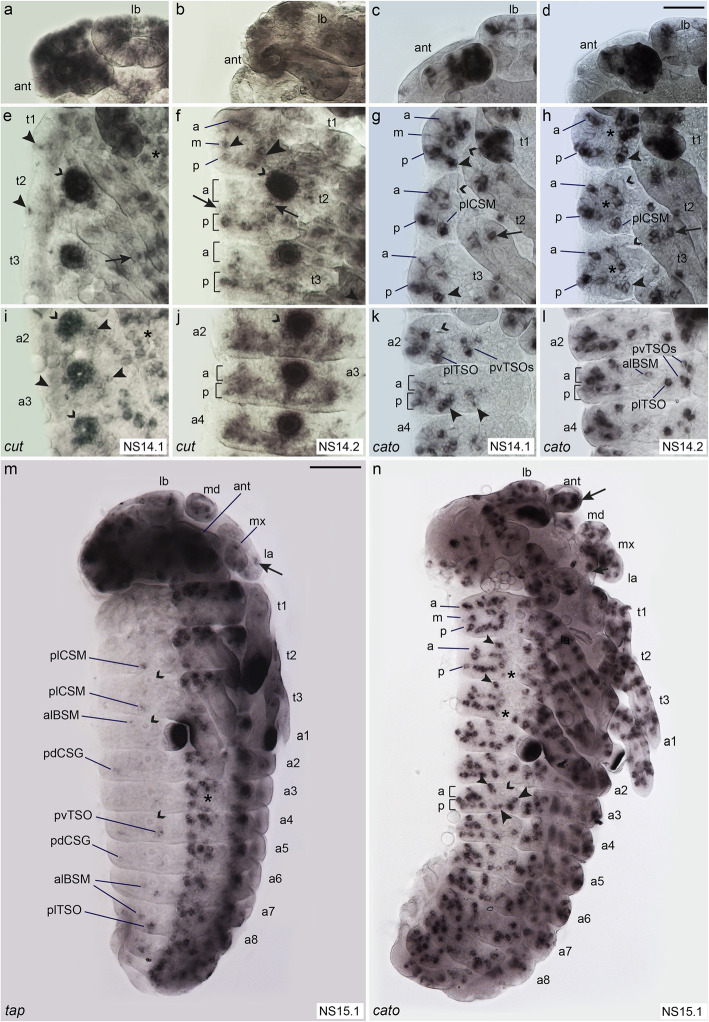
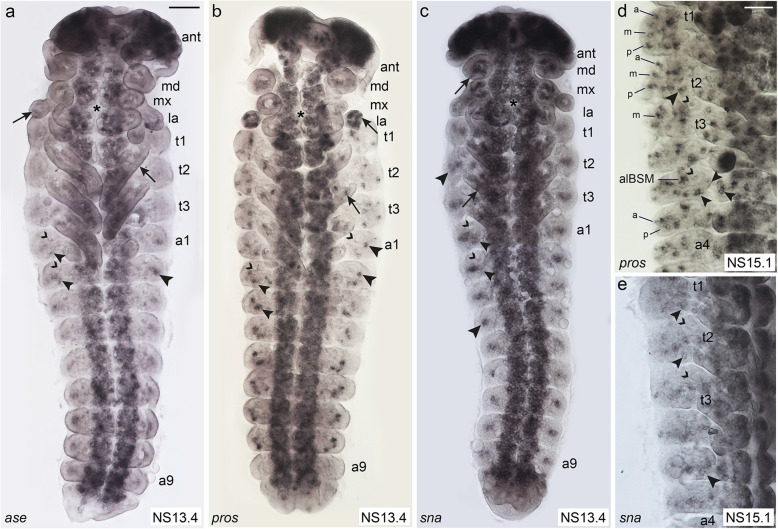
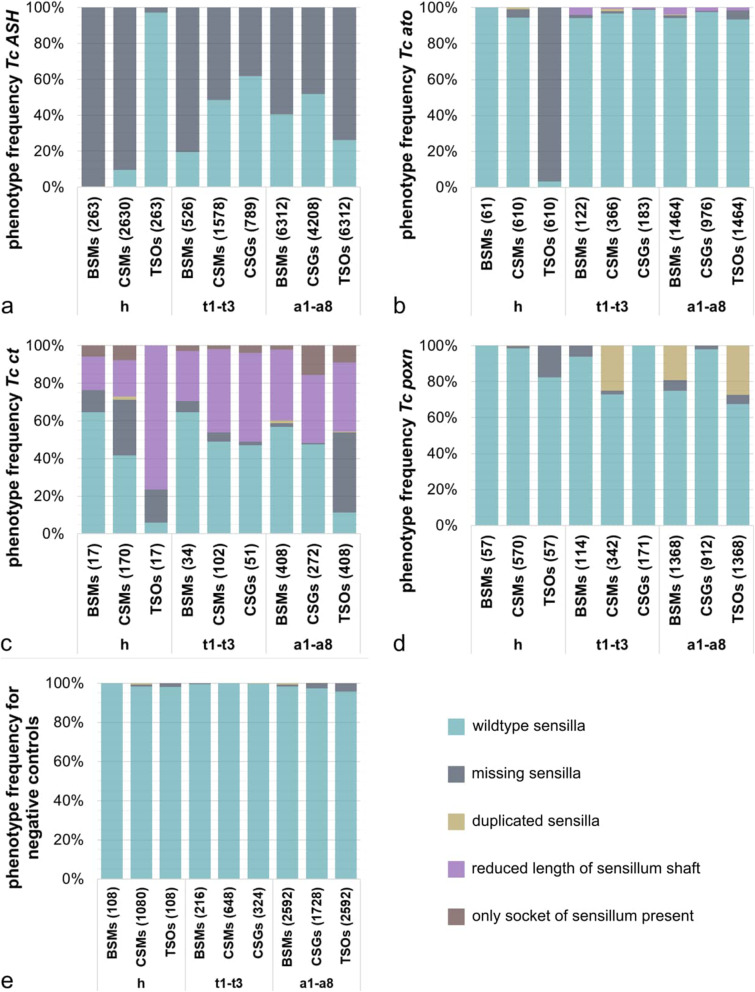
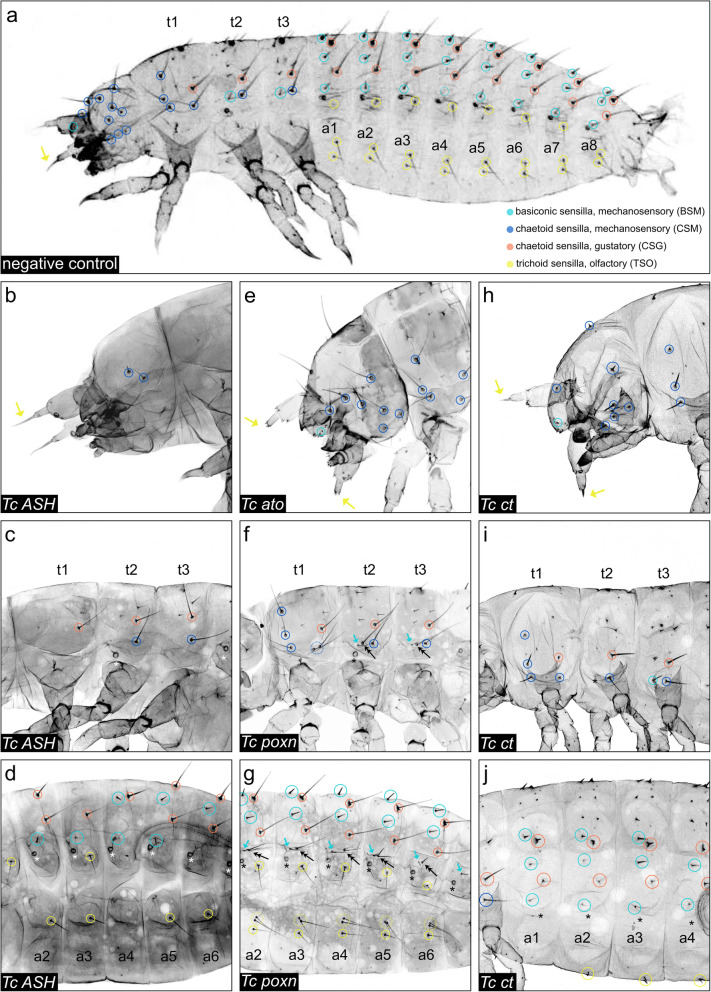
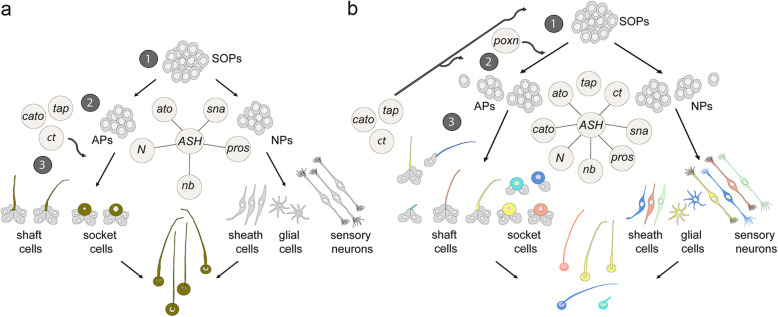
Insects and other arthropods utilise external sensory structures for mechanosensory, olfactory, and gustatory reception. These sense organs have characteristic shapes related to their function, and in many cases are distributed in a fixed pattern so that they are identifiable individually. In Drosophila melanogaster, the identity of sense organs is regulated by specific combinations of transcription factors. In other arthropods, however, sense organ subtypes cannot be linked to the same code of gene expression. This raises the questions of how sense organ diversity has evolved and whether the principles underlying subtype identity in D. melanogaster are representative of other insects. Here, we provide evidence that such principles cannot be generalised, and suggest that sensory organ diversification followed the recruitment of sensory genes to distinct sensory organ specification mechanism. RESULTS: We analysed sense organ development in a nondipteran insect, the flour beetle Tribolium castaneum, by gene expression and RNA interference studies. We show that in contrast to D. melanogaster, T. castaneum sense organs cannot be categorised based on the expression or their requirement for individual or combinations of conserved sense organ transcription factors such as cut and pox neuro, or members of the Achaete-Scute (Tc ASH, Tc asense), Atonal (Tc atonal, Tc cato, Tc amos), and neurogenin families (Tc tap). Rather, our observations support an evolutionary scenario whereby these sensory genes are required for the specification of sense organ precursors and the development and differentiation of sensory cell types in diverse external sensilla which do not fall into specific morphological and functional classes. CONCLUSIONS: Based on our findings and past research, we present an evolutionary scenario suggesting that sense organ subtype identity has evolved by recruitment of a flexible sensory gene network to the different sense organ specification processes. A dominant role of these genes in subtype identity has evolved as a secondary effect of the function of these genes in individual or subsets of sense organs, probably modulated by positional cues.
Keywords: Evolution; Gene expression; RNA interference; Sense organ development; Sense organ subtypes; Tribolium castaneum.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures

Similar articles
-
Hox genes require homothorax and extradenticle for body wall identity specification but not for appendage identity specification during metamorphosis of Tribolium castaneum.Dev Biol. 2014 Nov 1;395(1):182-97. doi: 10.1016/j.ydbio.2014.08.017. Epub 2014 Sep 4. Dev Biol. 2014. PMID: 25195194
-
The expression and function of the achaete-scute genes in Tribolium castaneum reveals conservation and variation in neural pattern formation and cell fate specification.Development. 2003 Sep;130(18):4373-81. doi: 10.1242/dev.00646. Development. 2003. PMID: 12900453
-
Evolutionary variation in neural gene expression in the developing sense organs of the crustacean Daphnia magna.Dev Biol. 2017 Apr 1;424(1):50-61. doi: 10.1016/j.ydbio.2017.02.011. Epub 2017 Feb 24. Dev Biol. 2017. PMID: 28238736
-
The specification of sensory neuron identity in Drosophila.Bioessays. 1993 May;15(5):293-8. doi: 10.1002/bies.950150502. Bioessays. 1993. PMID: 8343140 Review.
-
A genome-wide inventory of neurohormone GPCRs in the red flour beetle Tribolium castaneum.Front Neuroendocrinol. 2008 Jan;29(1):142-65. doi: 10.1016/j.yfrne.2007.10.003. Epub 2007 Oct 24. Front Neuroendocrinol. 2008. PMID: 18054377 Review.
Cited by
-
Probing the conserved roles of cut in the development and function of optically different insect compound eyes.Front Cell Dev Biol. 2023 Mar 31;11:1104620. doi: 10.3389/fcell.2023.1104620. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 37065850 Free PMC article.
-
Lepidopteran scale cells derive from sensory organ precursors through a canonical lineage.Development. 2025 Mar 1;152(5):DEV204501. doi: 10.1242/dev.204501. Epub 2025 Mar 7. Development. 2025. PMID: 40052482 Free PMC article.
References
-
- Barth FG. A spider’s world - senses and behavior: Springer; 2001. www.springer.com/de/book/9783540420460. Accessed 20 Nov 2017
-
- Stevens M. Sensory ecology, behaviour, and evolution. Oxford: Oxford University Press; 2013.
-
- Shanbhag SR, Müller B, Steinbrecht RA. Atlas of olfactory organs of Drosophila melanogaster: 1. Types, external organization, innervation and distribution of olfactory sensilla. Int J Insect Morph Embryol. 1999;28:377–397. doi: 10.1016/S0020-7322(99)00039-2. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
