

Fairy circles reveal the resilience of self-organized salt marshes
- PMID: 33547078
- PMCID: PMC7864568
- DOI: 10.1126/sciadv.abe1100
Fairy circles reveal the resilience of self-organized salt marshes
Abstract
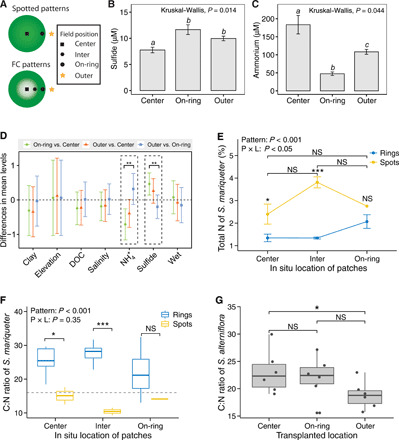
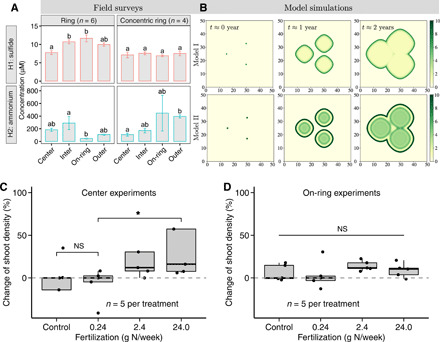
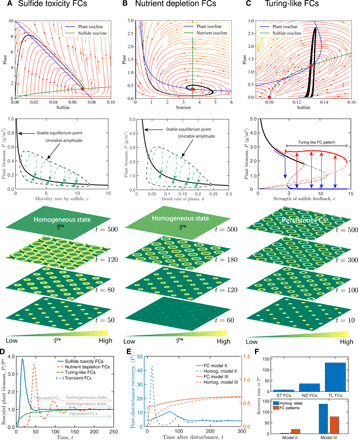
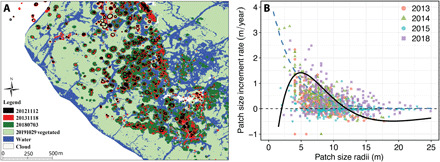
Spatial patterning is a fascinating theme in both theoretical and experimental ecology. It reveals resilience and stability to withstand external disturbances and environmental stresses. However, existing studies mainly focus on well-developed persistent patterns rather than transient patterns in self-organizing ecosystems. Here, combining models and experimental evidence, we show that transient fairy circle patterns in intertidal salt marshes can both infer the underlying ecological mechanisms and provide a measure of resilience. The models based on sulfide accumulation and nutrient depletion mechanisms reproduced the field-observed fairy circles, providing a generalized perspective on the emergence of transient patterns in salt marsh ecosystems. Field experiments showed that nitrogen fertilization mitigates depletion stress and shifts plant growth from negative to positive in the center of patches. Hence, nutrient depletion plays an overriding role, as only this process can explain the concentric rings. Our findings imply that the emergence of transient patterns can identify the ecological processes underlying pattern formation and the factors determining the ecological resilience of salt marsh ecosystems.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures

References
-
- Klausmeier C. A., Regular and irregular patterns in semiarid vegetation. Science 284, 1826–1828 (1999). - PubMed
-
- Rietkerk M., Dekker S. C., de Ruiter P. C., van de Koppel J., Self-organized patchiness and catastrophic shifts in ecosystems. Science 305, 1926–1929 (2004). - PubMed
-
- Pringle R. M., Tarnita C. E., Spatial self-organization of ecosystems: Integrating multiple mechanisms of regular-pattern formation. Annu. Rev. Entomol. 62, 359–377 (2017). - PubMed
-
- van de Koppel J., Gascoigne J. C., Therautaz G., Rietkerk M., Mooij W. M., Herman P. M. J., Experimental evidence for spatial self-organization and its emergent effects in mussel bed ecosystems. Science 322, 739–742 (2008). - PubMed
-
- van de Koppel J., Rietkerk M., Dankers N., Herman P. M. J., Scale-dependent feedback and regular spatial patterns in young mussel beds. Am. Nat. 165, E66–E77 (2005). - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
