

Nsp1 protein of SARS-CoV-2 disrupts the mRNA export machinery to inhibit host gene expression
- PMID: 33547084
- PMCID: PMC7864571
- DOI: 10.1126/sciadv.abe7386
Nsp1 protein of SARS-CoV-2 disrupts the mRNA export machinery to inhibit host gene expression
Abstract
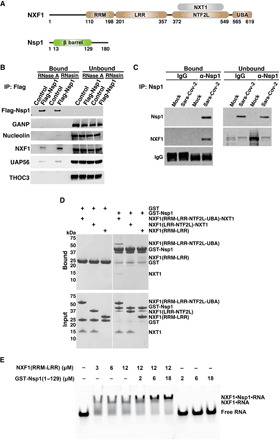
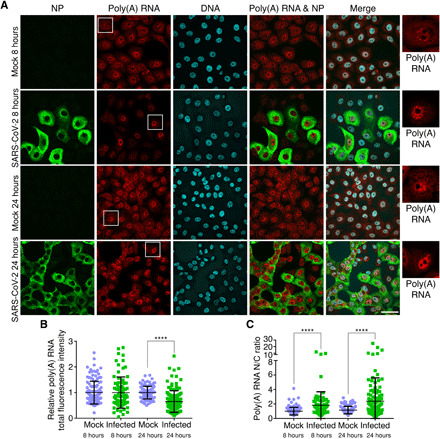
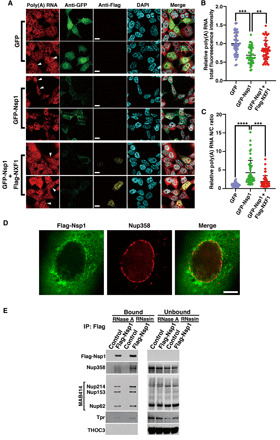
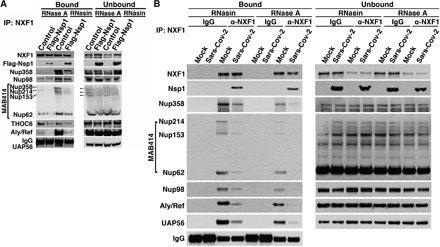
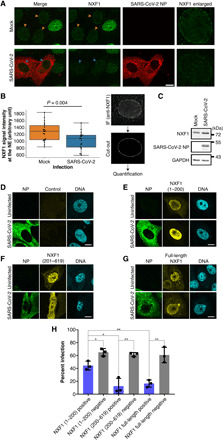
The ongoing unprecedented severe acute respiratory syndrome caused by the SARS-CoV-2 outbreak worldwide has highlighted the need for understanding viral-host interactions involved in mechanisms of virulence. Here, we show that the virulence factor Nsp1 protein of SARS-CoV-2 interacts with the host messenger RNA (mRNA) export receptor heterodimer NXF1-NXT1, which is responsible for nuclear export of cellular mRNAs. Nsp1 prevents proper binding of NXF1 to mRNA export adaptors and NXF1 docking at the nuclear pore complex. As a result, a significant number of cellular mRNAs are retained in the nucleus during infection. Increased levels of NXF1 rescues the Nsp1-mediated mRNA export block and inhibits SARS-CoV-2 infection. Thus, antagonizing the Nsp1 inhibitory function on mRNA export may represent a strategy to restoring proper antiviral host gene expression in infected cells.
Copyright © 2021 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution License 4.0 (CC BY).
Figures
Comment in
-
Attenuating innate immunity and facilitating β-coronavirus infection by NSP1 of SARS-CoV-2 through specific redistributing hnRNP A2/B1 cellular localization.Signal Transduct Target Ther. 2021 Oct 26;6(1):371. doi: 10.1038/s41392-021-00786-y. Signal Transduct Target Ther. 2021. PMID: 34702797 Free PMC article. No abstract available.
References
-
- Ayllon J., García-Sastre A., The NS1 protein: A multitasking virulence factor. Curr. Top. Microbiol. Immunol. 386, 73–107 (2015). - PubMed
-
- Zhang L., Das P., Schmolke M., Manicassamy B., Wang Y., Deng X., Cai L., Tu B. P., Forst C. V., Roth M. G., Levy D. E., García-Sastre A., de Brabander J., Phillips M. A., Fontoura B. M. A., Inhibition of pyrimidine synthesis reverses viral virulence factor-mediated block of mRNA nuclear export. J. Cell Biol. 196, 315–326 (2012). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
