

Precision of tissue patterning is controlled by dynamical properties of gene regulatory networks
- PMID: 33547135
- PMCID: PMC7929933
- DOI: 10.1242/dev.197566
Precision of tissue patterning is controlled by dynamical properties of gene regulatory networks
Abstract
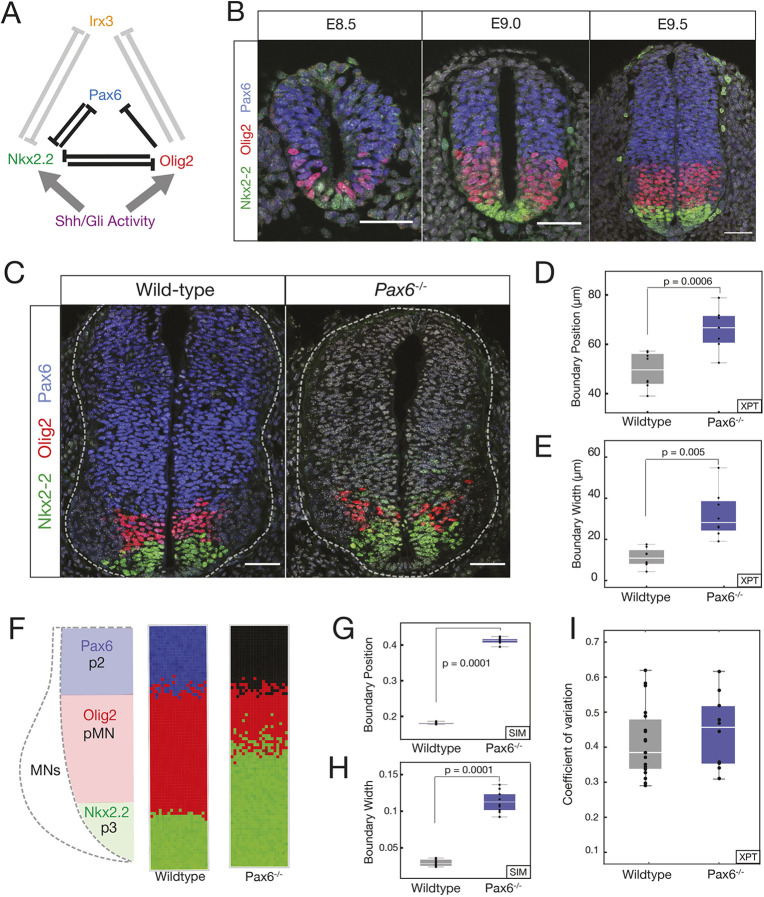
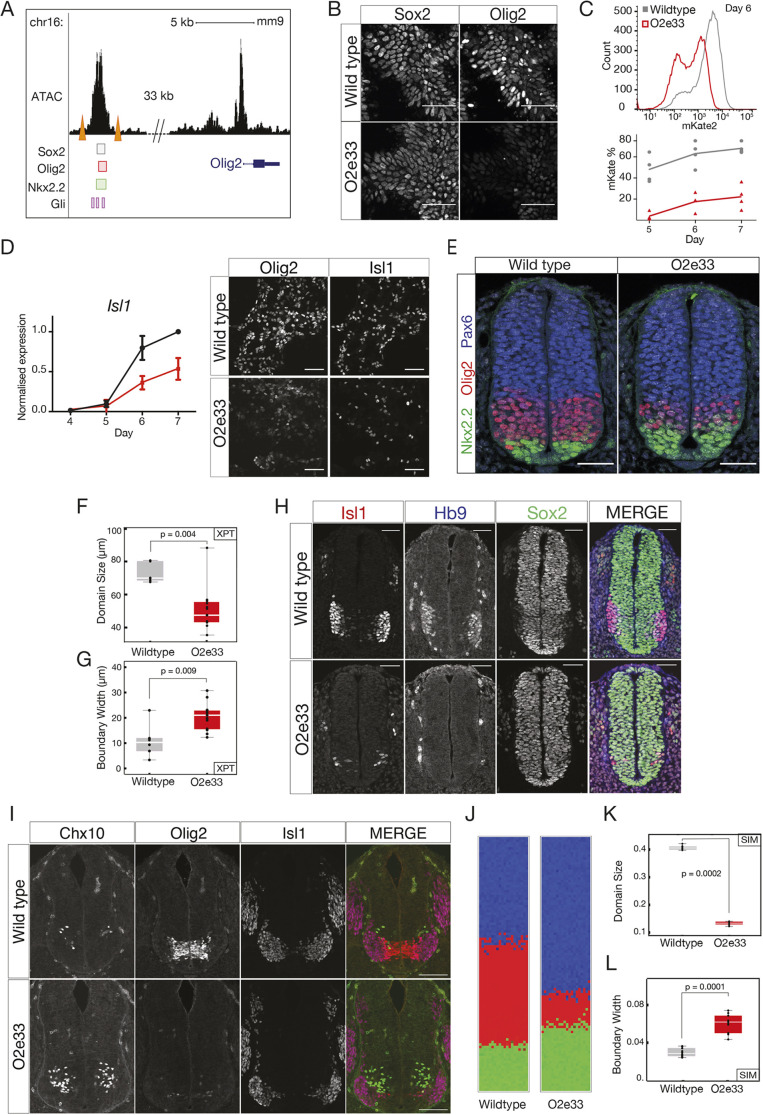
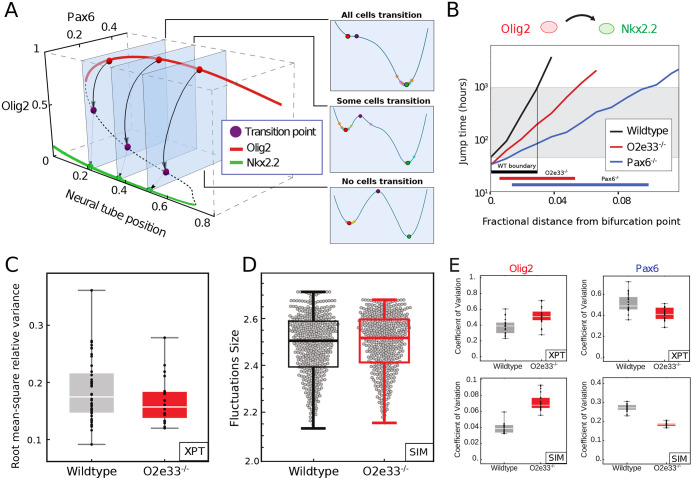
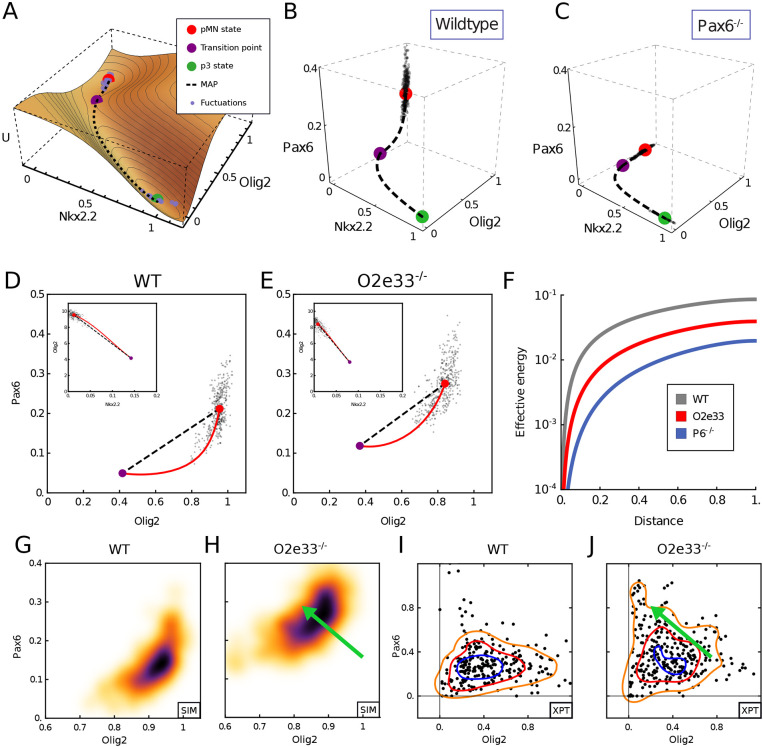
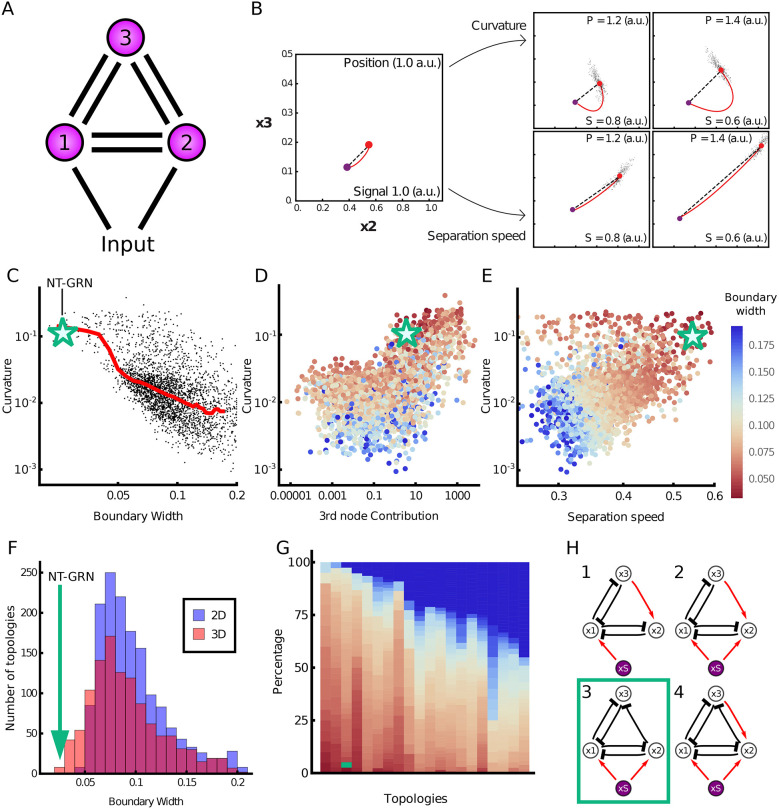
During development, gene regulatory networks allocate cell fates by partitioning tissues into spatially organised domains of gene expression. How the sharp boundaries that delineate these gene expression patterns arise, despite the stochasticity associated with gene regulation, is poorly understood. We show, in the vertebrate neural tube, using perturbations of coding and regulatory regions, that the structure of the regulatory network contributes to boundary precision. This is achieved, not by reducing noise in individual genes, but by the configuration of the network modulating the ability of stochastic fluctuations to initiate gene expression changes. We use a computational screen to identify network properties that influence boundary precision, revealing two dynamical mechanisms by which small gene circuits attenuate the effect of noise in order to increase patterning precision. These results highlight design principles of gene regulatory networks that produce precise patterns of gene expression.
Keywords: Dynamical systems theory; Gene regulatory network; Morphogen signaling; Neural tube; cis regulatory elements.
© 2021. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interestsThe authors declare no competing or financial interests.
Figures
References
-
- Akam, M. (1987). The molecular basis for metameric pattern in the Drosophila embryo. Development 101, 1-22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
