

Extracellular DNA, cell surface proteins and c-di-GMP promote biofilm formation in Clostridioides difficile
- PMID: 33547340
- PMCID: PMC7865049
- DOI: 10.1038/s41598-020-78437-5
Extracellular DNA, cell surface proteins and c-di-GMP promote biofilm formation in Clostridioides difficile
Abstract
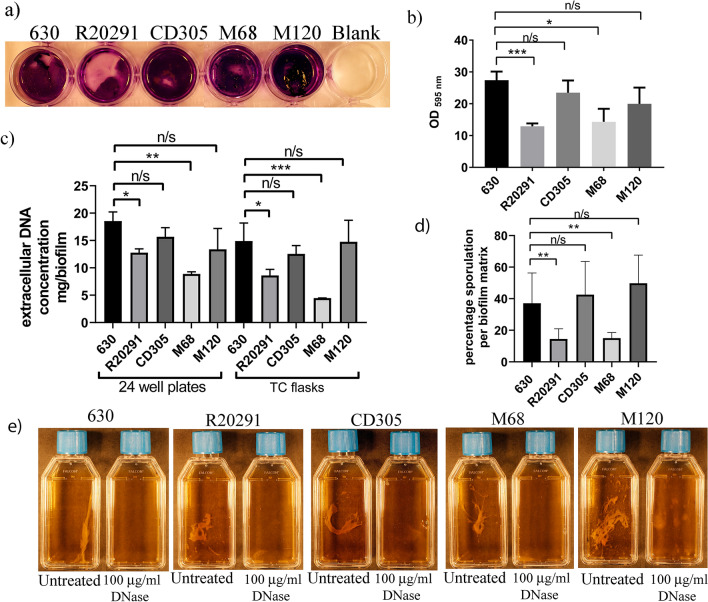
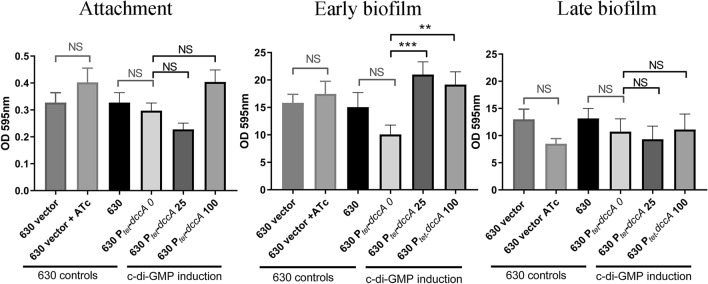
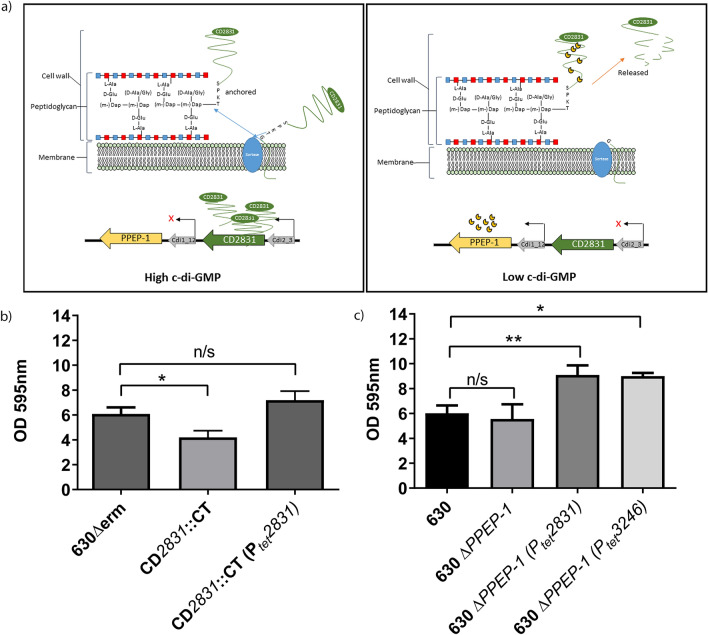
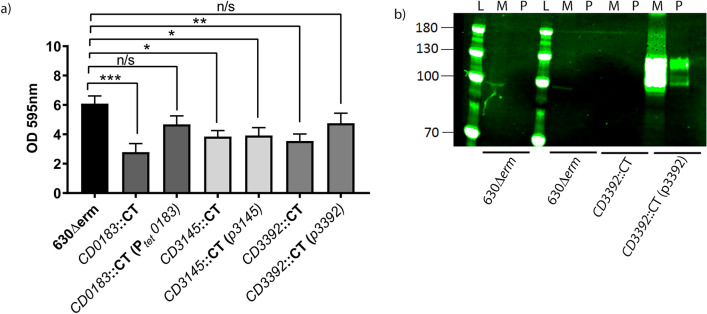
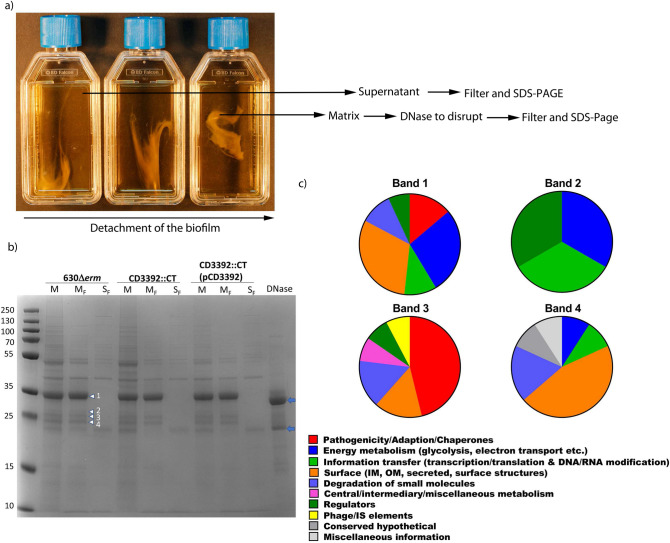
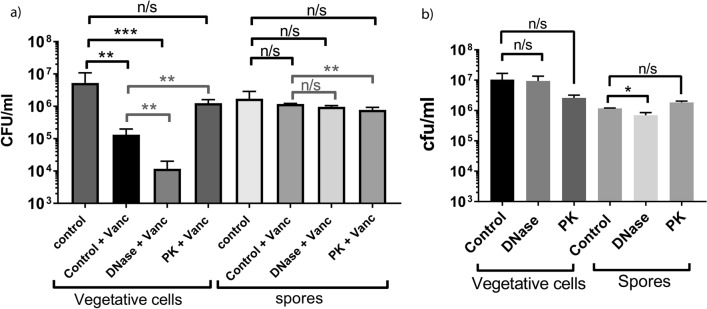
Clostridioides difficile is the leading cause of nosocomial antibiotic-associated diarrhoea worldwide, yet there is little insight into intestinal tract colonisation and relapse. In many bacterial species, the secondary messenger cyclic-di-GMP mediates switching between planktonic phase, sessile growth and biofilm formation. We demonstrate that c-di-GMP promotes early biofilm formation in C. difficile and that four cell surface proteins contribute to biofilm formation, including two c-di-GMP regulated; CD2831 and CD3246, and two c-di-GMP-independent; CD3392 and CD0183. We demonstrate that C. difficile biofilms are composed of extracellular DNA (eDNA), cell surface and intracellular proteins, which form a protective matrix around C. difficile vegetative cells and spores, as shown by a protective effect against the antibiotic vancomycin. We demonstrate a positive correlation between biofilm biomass, sporulation frequency and eDNA abundance in all five C. difficile lineages. Strains 630 (RT012), CD305 (RT023) and M120 (RT078) contain significantly more eDNA in their biofilm matrix than strains R20291 (RT027) and M68 (RT017). DNase has a profound effect on biofilm integrity, resulting in complete disassembly of the biofilm matrix, inhibition of biofilm formation and reduced spore germination. The addition of exogenous DNase could be exploited in treatment of C. difficile infection and relapse, to improve antibiotic efficacy.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
c-di-GMP Inhibits Early Sporulation in Clostridioides difficile.mSphere. 2021 Dec 22;6(6):e0091921. doi: 10.1128/msphere.00919-21. Epub 2021 Dec 8. mSphere. 2021. PMID: 34878288 Free PMC article.
-
Cyclic diguanylate differentially regulates the expression of virulence factors and pathogenesis-related phenotypes in Clostridioides difficile.Microbiol Res. 2024 Sep;286:127811. doi: 10.1016/j.micres.2024.127811. Epub 2024 Jun 20. Microbiol Res. 2024. PMID: 38909416 Review.
-
A Nutrient-Regulated Cyclic Diguanylate Phosphodiesterase Controls Clostridium difficile Biofilm and Toxin Production during Stationary Phase.Infect Immun. 2017 Aug 18;85(9):e00347-17. doi: 10.1128/IAI.00347-17. Print 2017 Sep. Infect Immun. 2017. PMID: 28652311 Free PMC article.
-
c-di-GMP turn-over in Clostridium difficile is controlled by a plethora of diguanylate cyclases and phosphodiesterases.PLoS Genet. 2011 Mar;7(3):e1002039. doi: 10.1371/journal.pgen.1002039. Epub 2011 Mar 31. PLoS Genet. 2011. PMID: 21483756 Free PMC article.
-
Biofilm regulation in Clostridioides difficile: Novel systems linked to hypervirulence.PLoS Pathog. 2021 Sep 9;17(9):e1009817. doi: 10.1371/journal.ppat.1009817. eCollection 2021 Sep. PLoS Pathog. 2021. PMID: 34499698 Free PMC article. Review.
Cited by
-
Clostridioides difficile Biofilm.Adv Exp Med Biol. 2024;1435:249-272. doi: 10.1007/978-3-031-42108-2_12. Adv Exp Med Biol. 2024. PMID: 38175479
-
Production of p-cresol by Decarboxylation of p-HPA by All Five Lineages of Clostridioides difficile Provides a Growth Advantage.Front Cell Infect Microbiol. 2021 Oct 29;11:757599. doi: 10.3389/fcimb.2021.757599. eCollection 2021. Front Cell Infect Microbiol. 2021. PMID: 34778108 Free PMC article.
-
c-di-GMP Inhibits Early Sporulation in Clostridioides difficile.mSphere. 2021 Dec 22;6(6):e0091921. doi: 10.1128/msphere.00919-21. Epub 2021 Dec 8. mSphere. 2021. PMID: 34878288 Free PMC article.
-
Revealing roles of S-layer protein (SlpA) in Clostridioides difficile pathogenicity by generating the first slpA gene deletion mutant.Microbiol Spectr. 2024 Jun 4;12(6):e0400523. doi: 10.1128/spectrum.04005-23. Epub 2024 May 6. Microbiol Spectr. 2024. PMID: 38709045 Free PMC article.
-
Rapid, Point-of-Care Microwave Lysis and Electrochemical Detection of Clostridioides difficile Directly from Stool Samples.Bioengineering (Basel). 2024 Jun 20;11(6):632. doi: 10.3390/bioengineering11060632. Bioengineering (Basel). 2024. PMID: 38927868 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
