

Planar cell polarity pathway in kidney development, function and disease
- PMID: 33547419
- PMCID: PMC8967065
- DOI: 10.1038/s41581-021-00395-6
Planar cell polarity pathway in kidney development, function and disease
Abstract
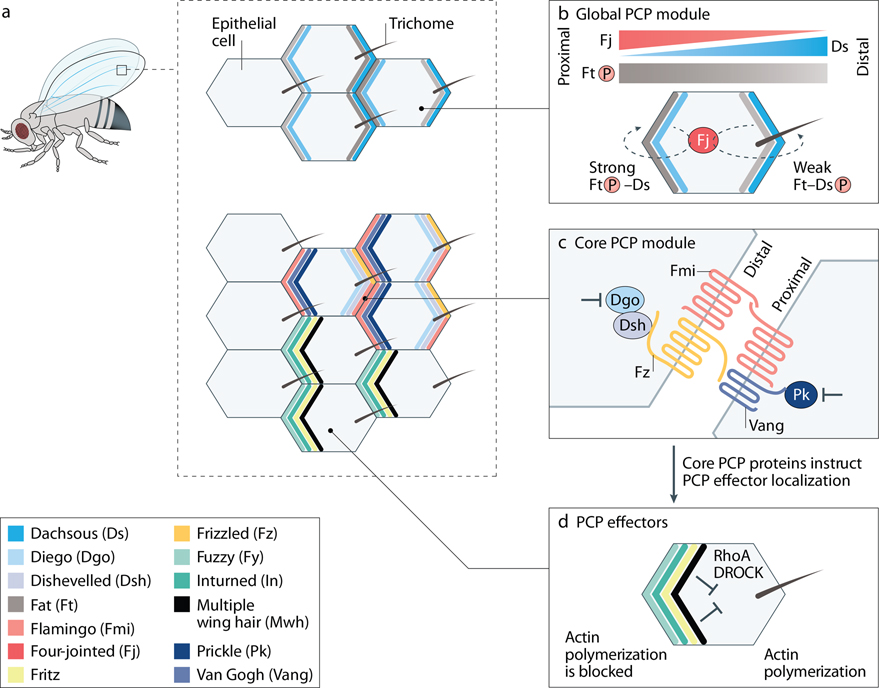
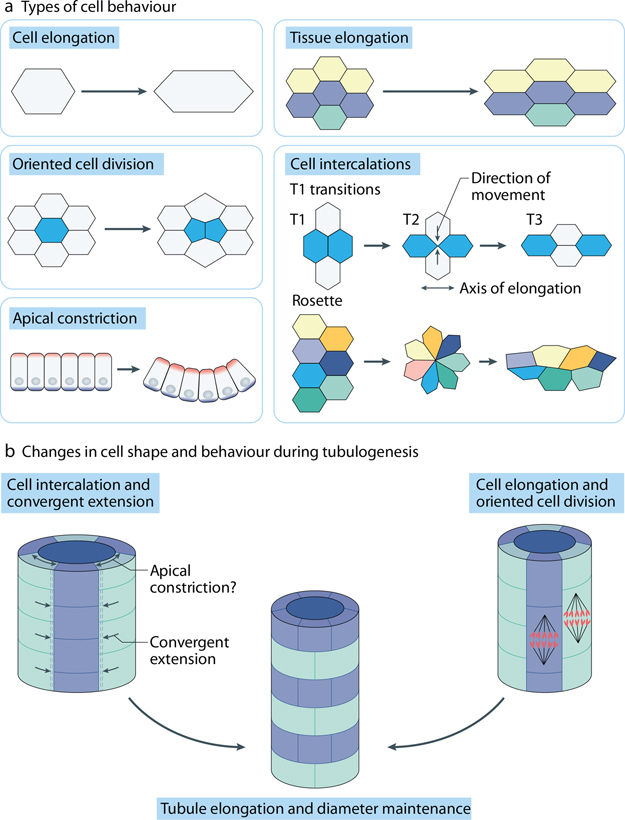
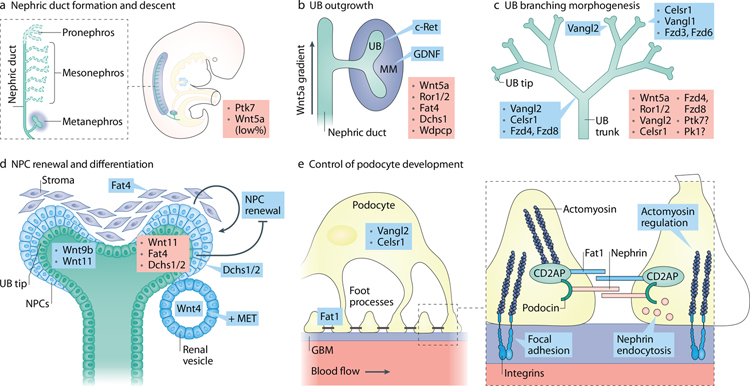
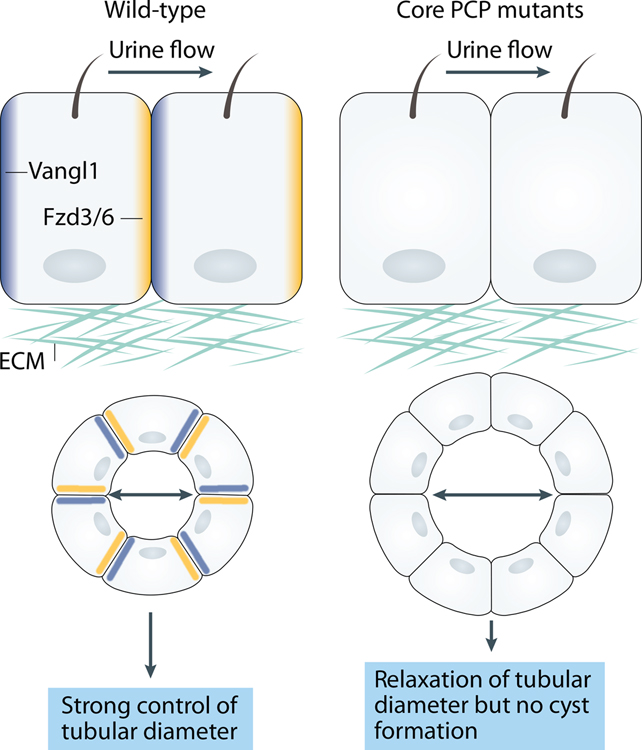
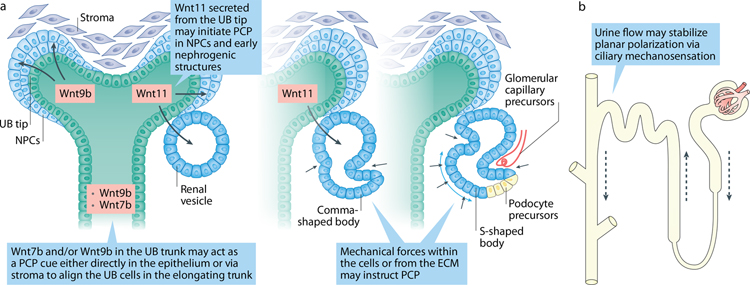
Planar cell polarity (PCP) refers to the coordinated orientation of cells in the tissue plane. Originally discovered and studied in Drosophila melanogaster, PCP is now widely recognized in vertebrates, where it is implicated in organogenesis. Specific sets of PCP genes have been identified. The proteins encoded by these genes become asymmetrically distributed to opposite sides of cells within a tissue plane and guide many processes that include changes in cell shape and polarity, collective cell movements or the uniform distribution of cell appendages. A unifying characteristic of these processes is that they often involve rearrangement of actomyosin. Mutations in PCP genes can cause malformations in organs of many animals, including humans. In the past decade, strong evidence has accumulated for a role of the PCP pathway in kidney development including outgrowth and branching morphogenesis of ureteric bud and podocyte development. Defective PCP signalling has been implicated in the pathogenesis of developmental kidney disorders of the congenital anomalies of the kidney and urinary tract spectrum. Understanding the origins, molecular constituents and cellular targets of PCP provides insights into the involvement of PCP molecules in normal kidney development and how dysfunction of PCP components may lead to kidney disease.
Figures
References
-
- Lawrence PA Gradients in the Insect Segment: The Orientation of Hairs in the Milkweed Bug Oncopeltus Fasciatus. Journal of Experimental Biology 44, 607–620 (1966). - PubMed
-
- Gubb D & Garcia-Bellido A A genetic analysis of the determination of cuticular polarity during development in Drosophila melanogaster. J Embryol Exp Morphol 68, 37–57 (1982). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
