

In vivo aggregation of presynaptic alpha-synuclein is not influenced by its phosphorylation at serine-129
- PMID: 33556542
- PMCID: PMC10405908
- DOI: 10.1016/j.nbd.2021.105291
In vivo aggregation of presynaptic alpha-synuclein is not influenced by its phosphorylation at serine-129
Abstract
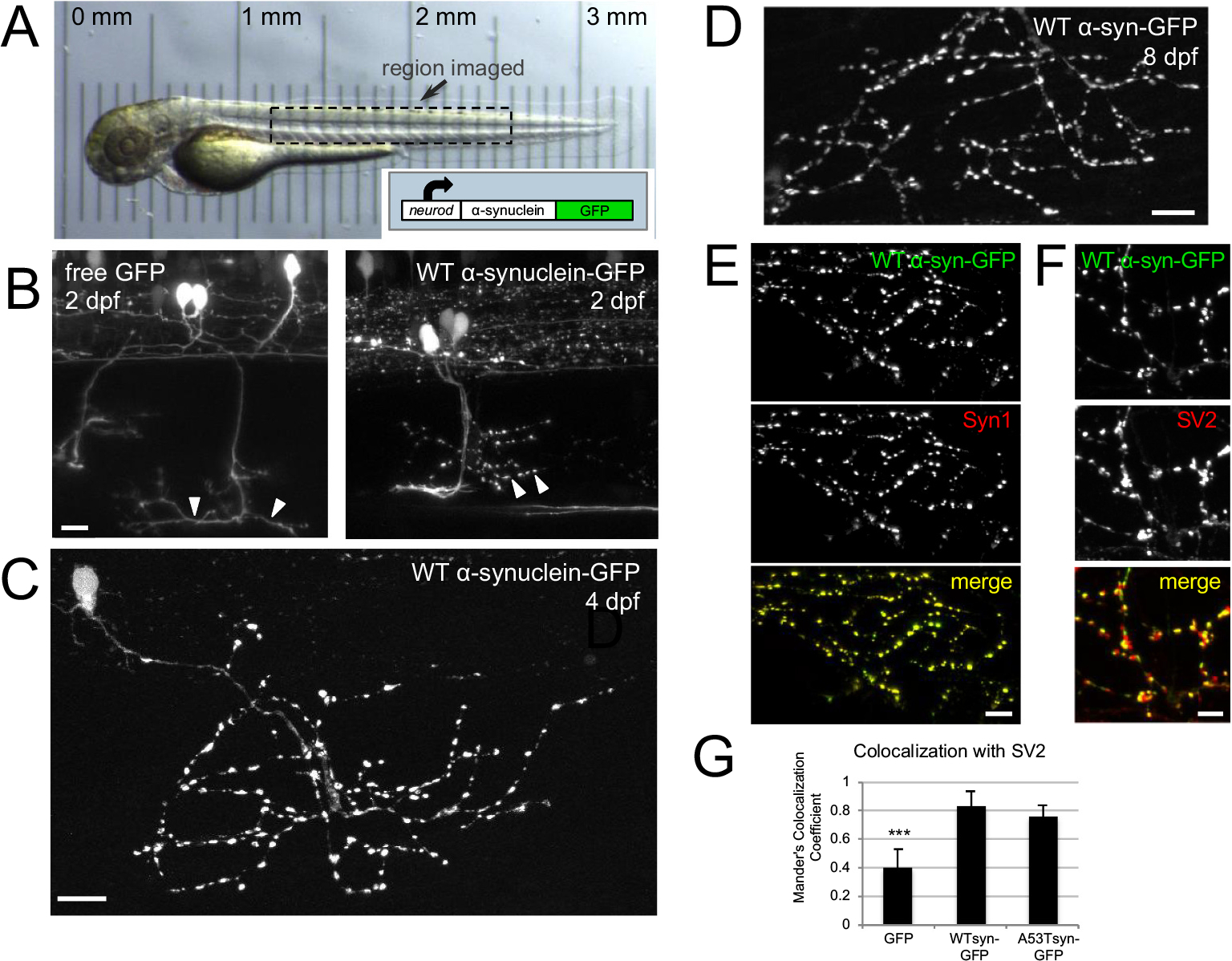
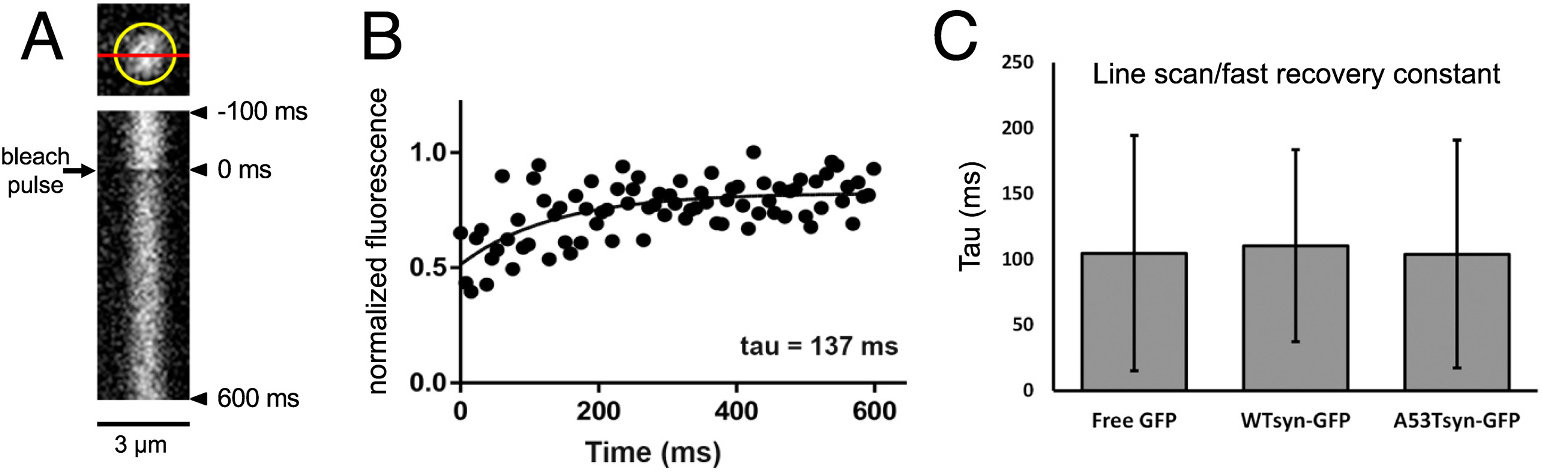
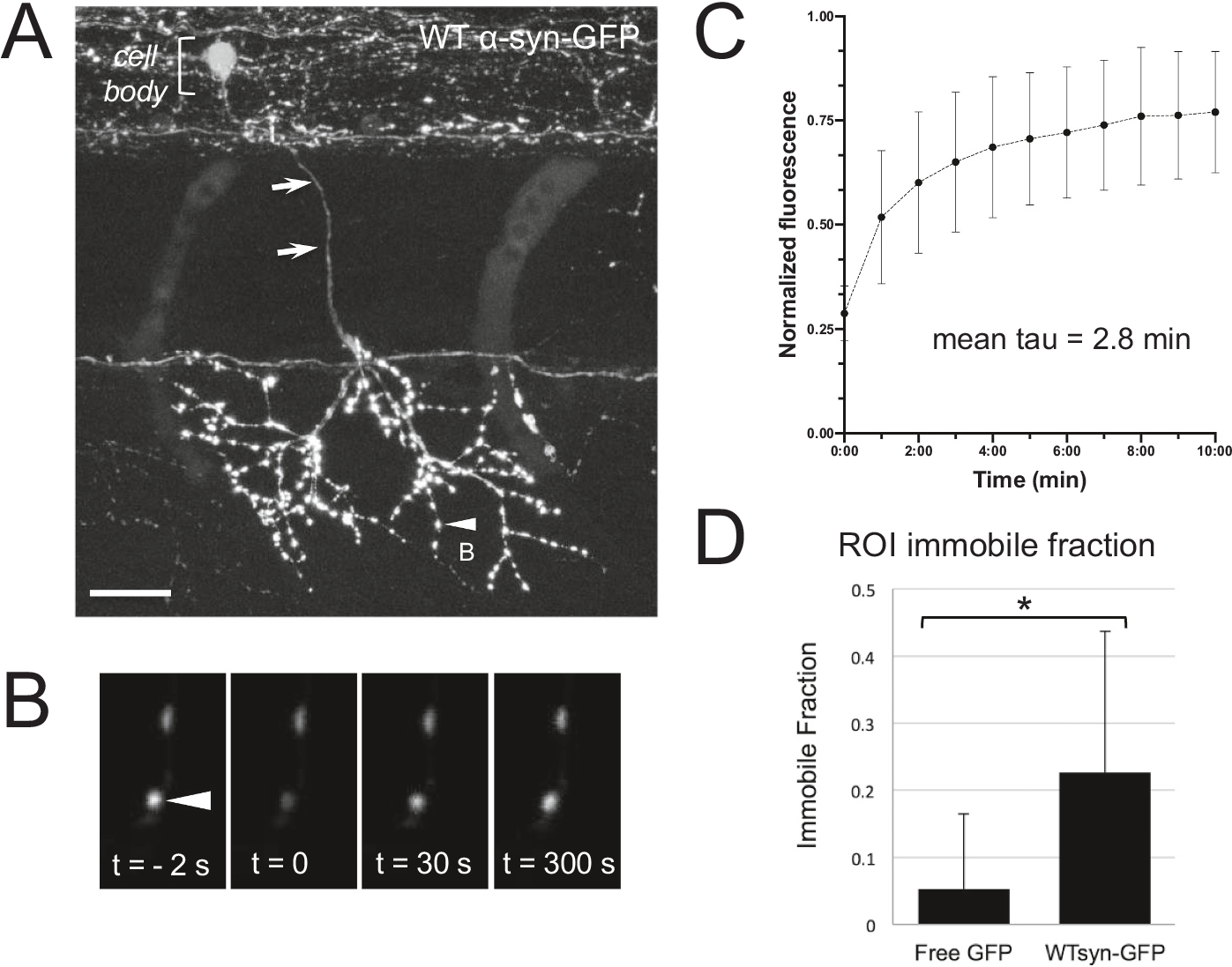
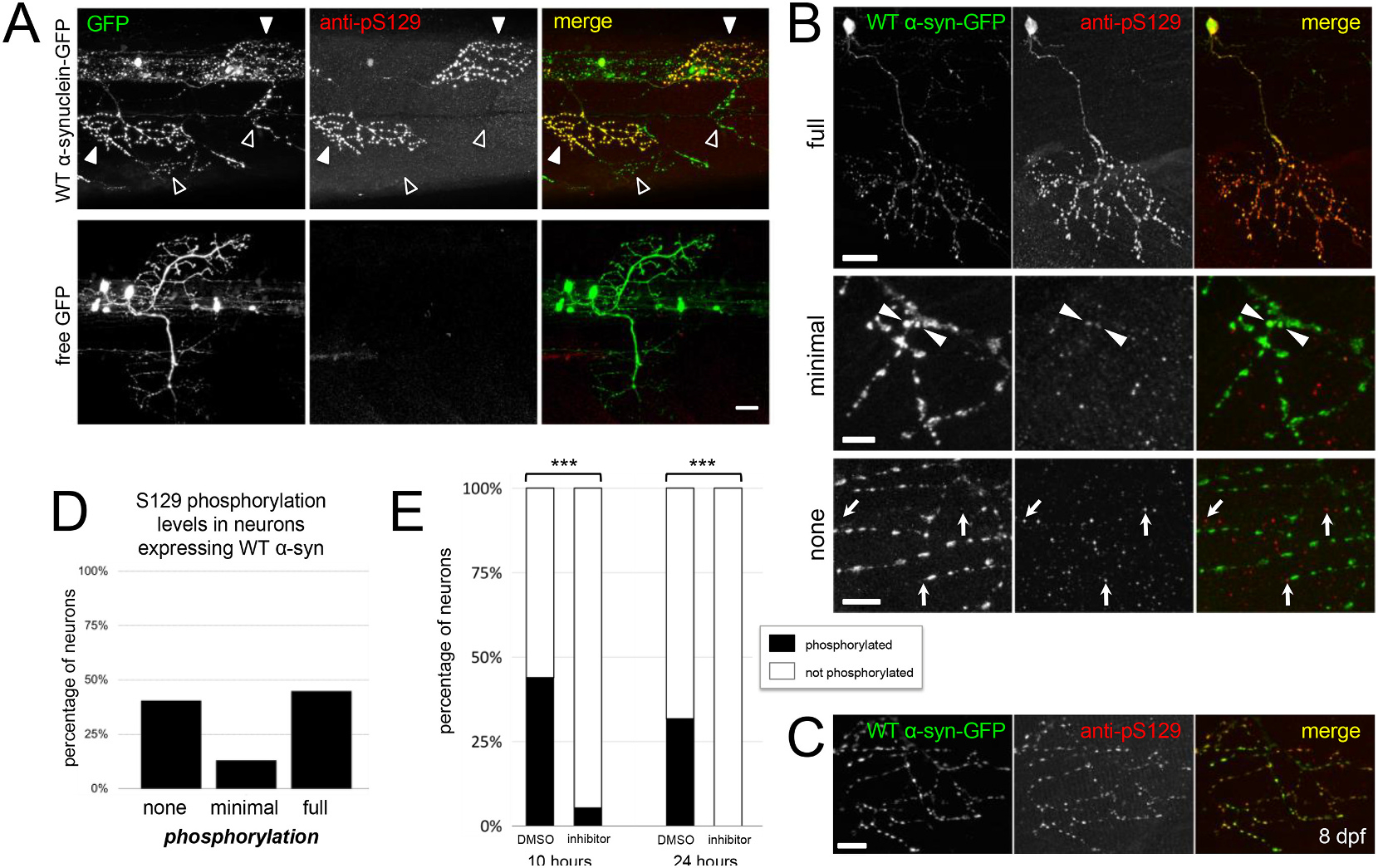
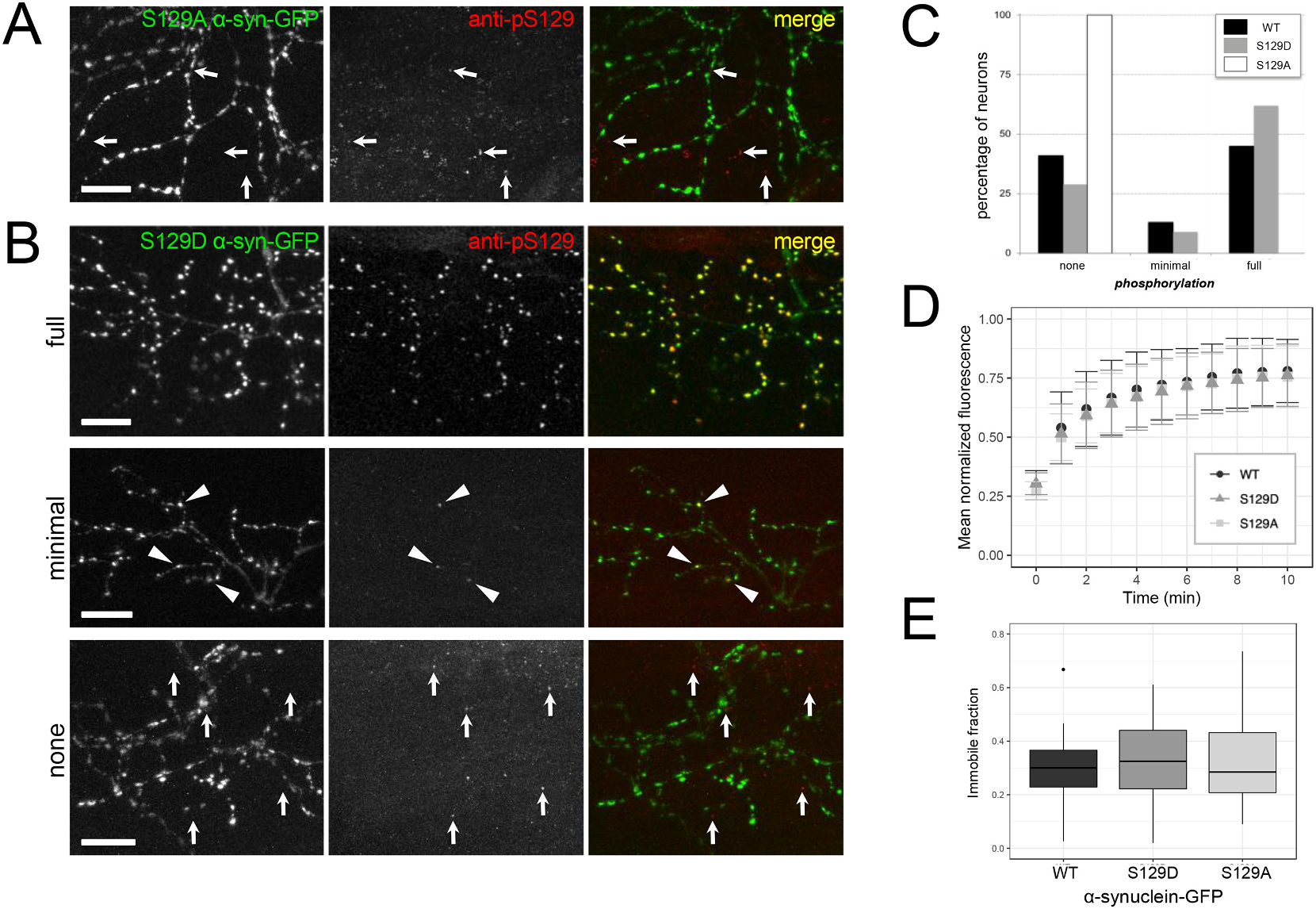
Abnormal aggregation of the α-synuclein protein is a key molecular feature of Parkinson's disease and other neurodegenerative diseases. The precise mechanisms that trigger α-synuclein aggregation are unclear, and it is not known what role aggregation plays in disease pathogenesis. Here we use an in vivo zebrafish model to express several different forms of human α-synuclein and measure its aggregation in presynaptic terminals. We show that human α-synuclein tagged with GFP can be expressed in zebrafish neurons, localizing normally to presynaptic terminals and undergoing phosphorylation at serine-129, as in mammalian neurons. The visual advantages of the zebrafish system allow for dynamic in vivo imaging to study α-synuclein, including the use of fluorescence recovery after photobleaching (FRAP) techniques to probe protein mobility. These experiments reveal three distinct terminal pools of α-synuclein with varying mobility, likely representing different subpopulations of aggregated and non-aggregated protein. Human α-synuclein is phosphorylated by an endogenous zebrafish Polo-like kinase activity, and there is a heterogeneous population of neurons containing either very little or extensive phosphorylation throughout the axonal arbor. Both pharmacological and genetic manipulations of serine-129 show that phosphorylation of α-synuclein at this site does not significantly affect its mobility. This suggests that serine-129 phosphorylation alone does not promote α-synuclein aggregation. Together our results show that human α-synuclein can be expressed and measured quantitatively in zebrafish, and that disease-relevant post-translational modifications occur within neurons. The zebrafish model provides a powerful in vivo system for measuring and manipulating α-synuclein function and aggregation, and for developing new treatments for neurodegenerative disease.
Keywords: Alpha-synuclein; FRAP; Parkinson's disease; Phosphorylation; Polo-like kinase; Protein aggregation; Zebrafish; in vivo imaging.
Copyright © 2021. Published by Elsevier Inc.
Figures
References
-
- Anderson JP, Walker DE, Goldstein JM, de Laat R, Banducci K, Caccavello RJ, Barbour R, Huang J, Kling K, Lee M, Diep L, Keim PS, Shen X, Chataway T, Schlossmacher MG, Seubert P, Schenk D, Sinha S, Gai WP, Chilcote TJ, 2006. Phosphorylation of Ser-129 is the dominant pathological modification of α-Synuclein in familial and sporadic Lewy body disease. J. Biol. Chem. 281, 29739–29752. - PubMed
-
- Bae Y-K, Kani S, Shimizu T, Tanabe K, Nojima H, Kimura Y, Higashijima S, Hibi M, 2009. Anatomy of zebrafish cerebellum and screen for mutations affecting its development. Dev. Biol. 330, 406–426. - PubMed
-
- Bergeron M, Motter R, Tanaka P, Fauss D, Babcock M, Chiou SS, Nelson S, San Pablo F, Anderson JP, 2014. In vivo modulation of polo-like kinases supports a key role for PLK2 in Ser129 α-synuclein phosphorylation in mouse brain. Neuroscience 256, 72–82. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
