

Spatiotemporal GLP-1 and GIP receptor signaling and trafficking/recycling dynamics induced by selected receptor mono- and dual-agonists
- PMID: 33556643
- PMCID: PMC7921015
- DOI: 10.1016/j.molmet.2021.101181
Spatiotemporal GLP-1 and GIP receptor signaling and trafficking/recycling dynamics induced by selected receptor mono- and dual-agonists
Abstract
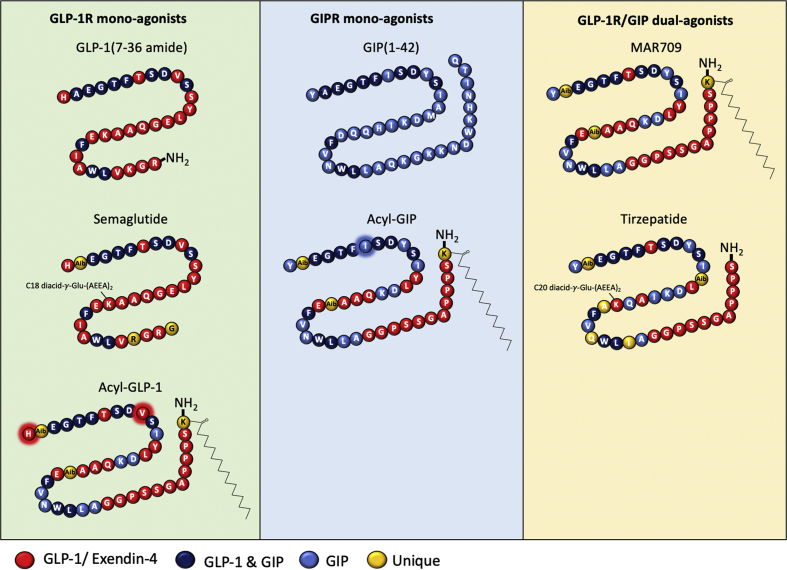
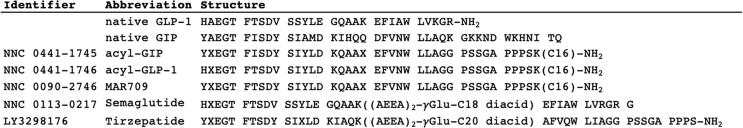
Objective: We assessed the spatiotemporal GLP-1 and GIP receptor signaling, trafficking, and recycling dynamics of GIPR mono-agonists, GLP-1R mono-agonists including semaglutide, and GLP-1/GIP dual-agonists MAR709 and tirzepatide.
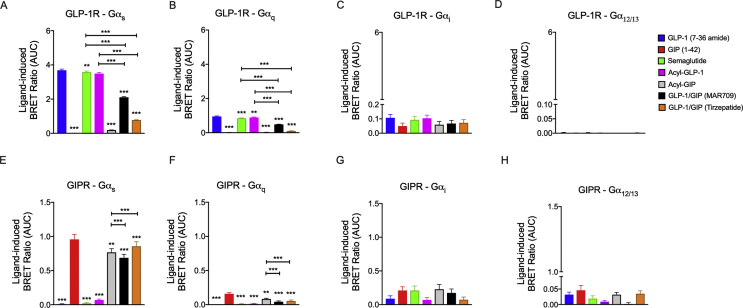
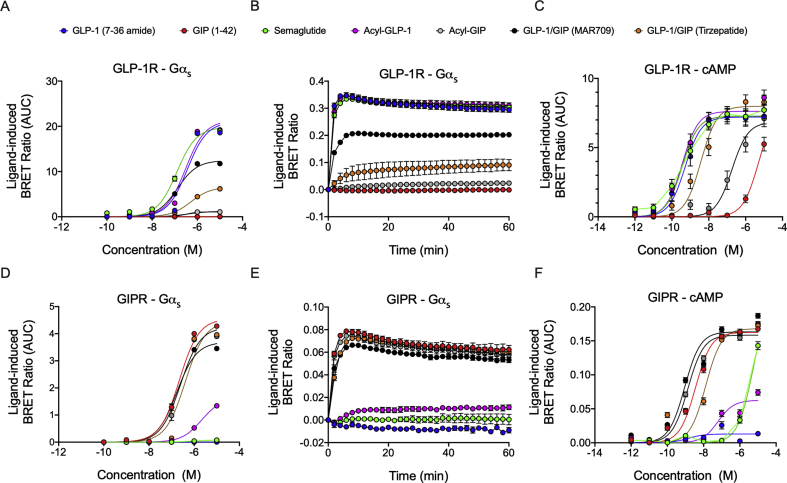
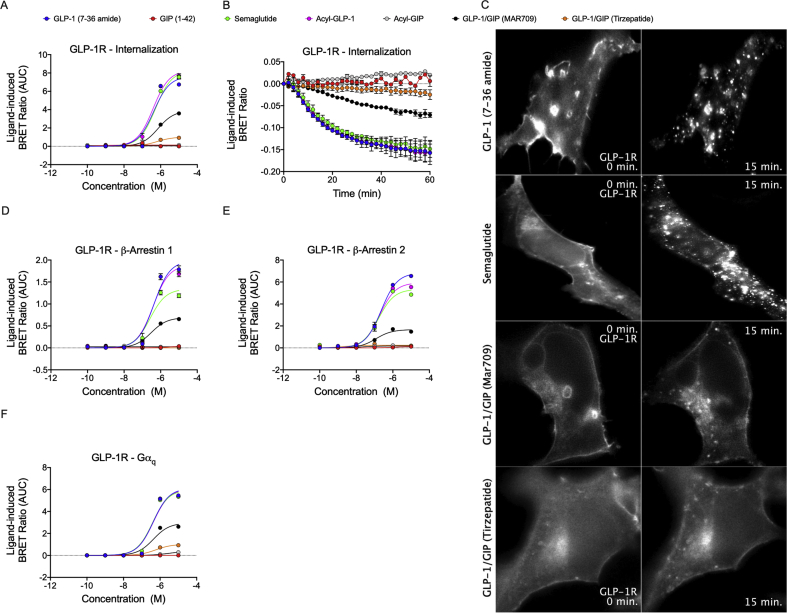
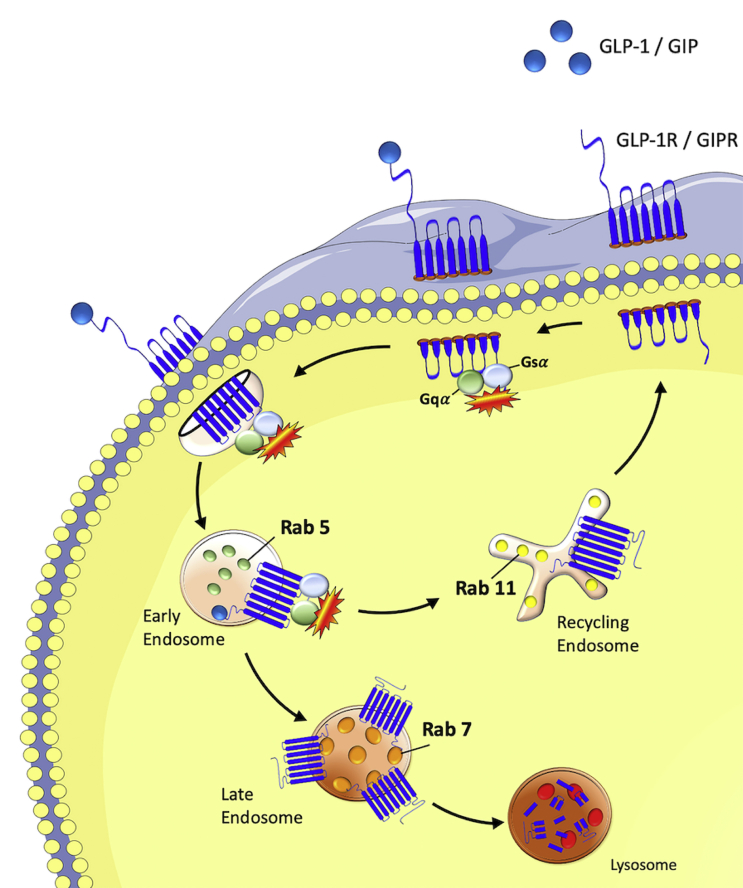
Methods: Receptor G protein recruitment and internalization/trafficking dynamics were assessed using bioluminescence resonance energy transfer (BRET)-based technology and live-cell HILO microscopy.
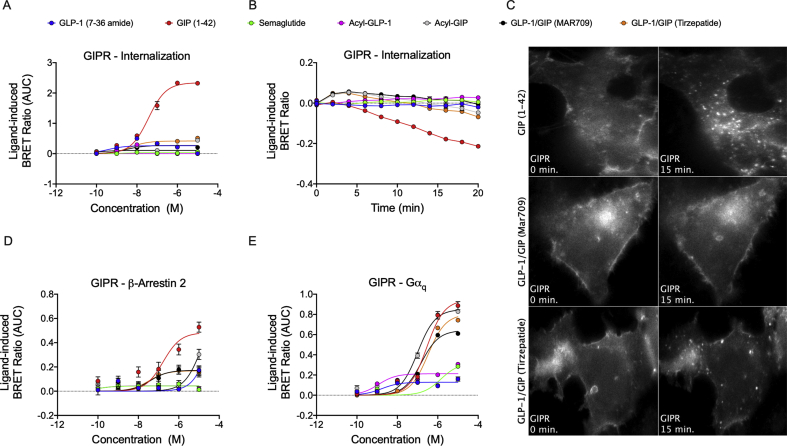
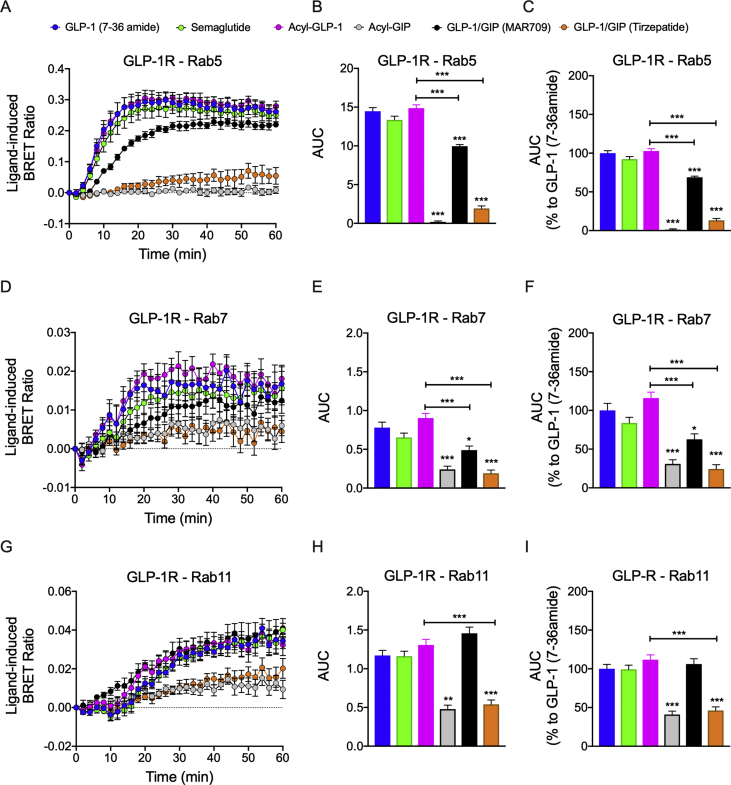
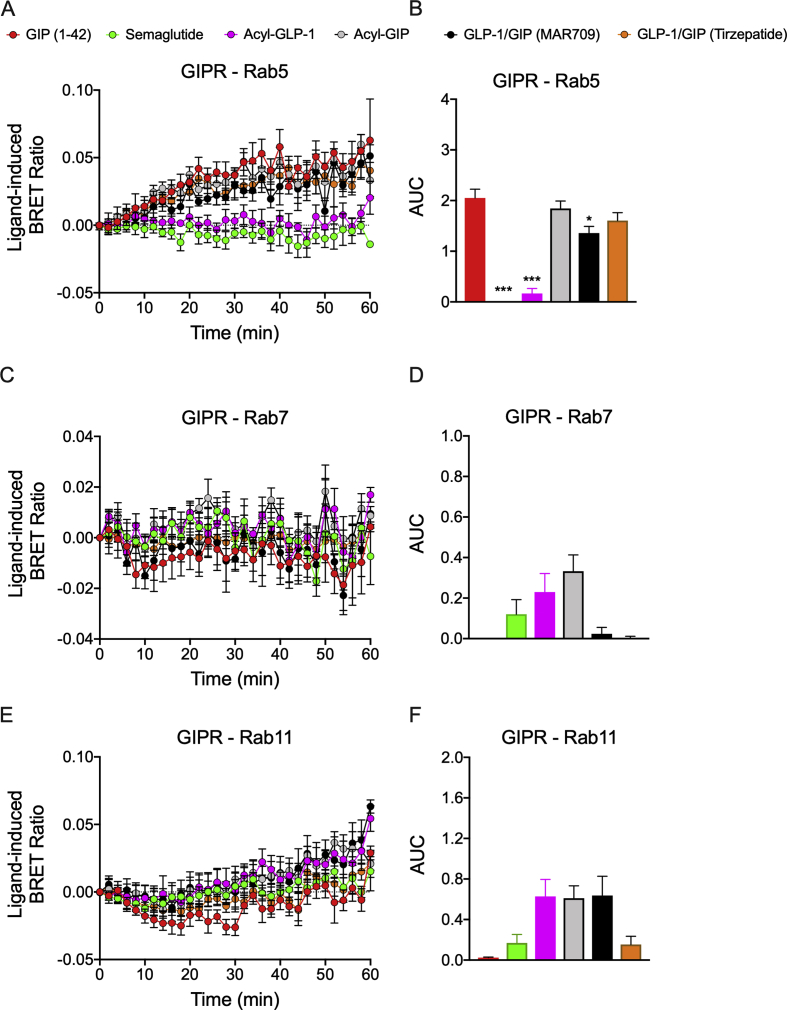
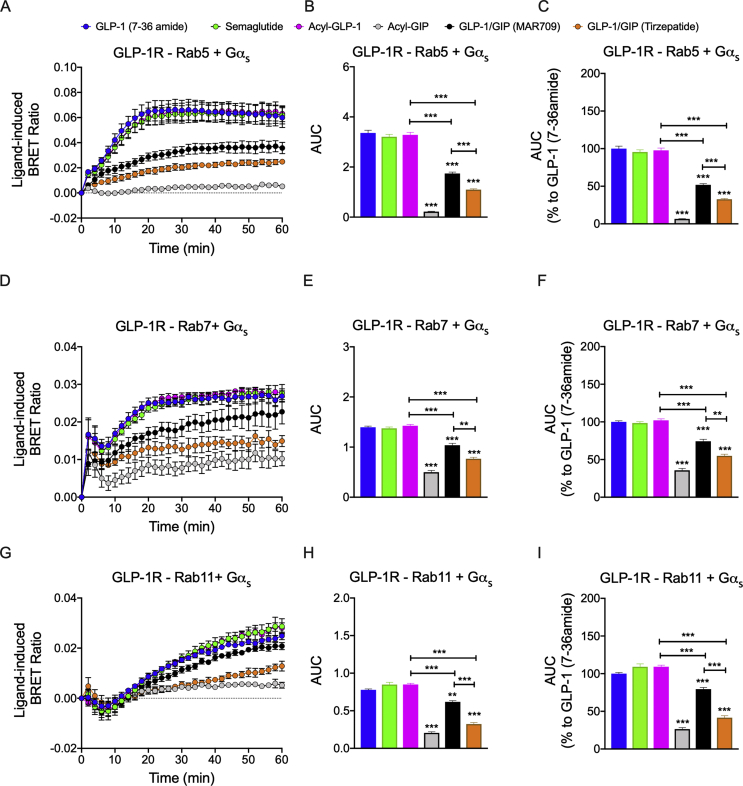
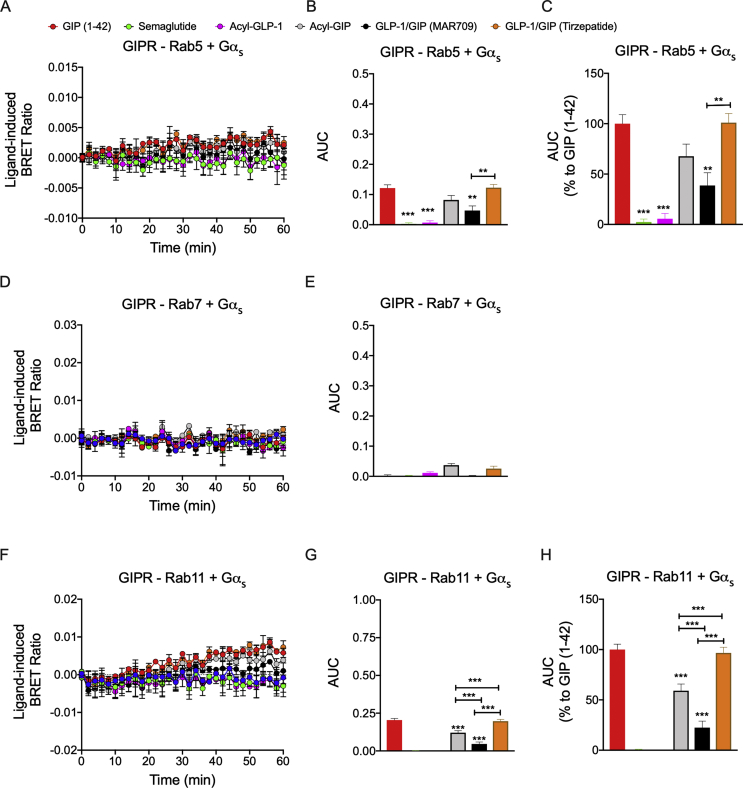
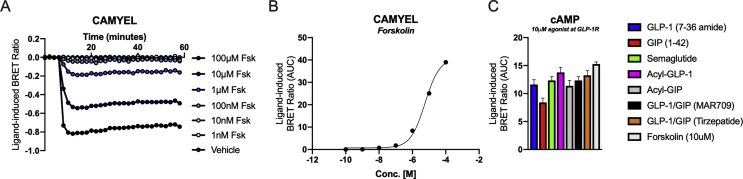
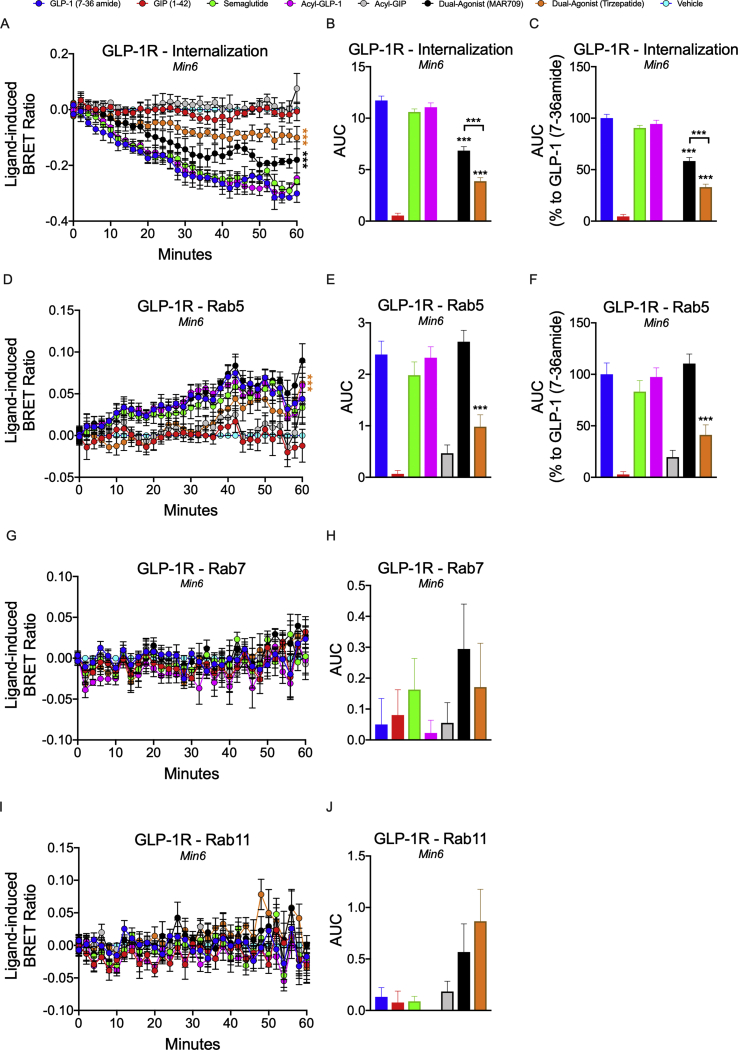
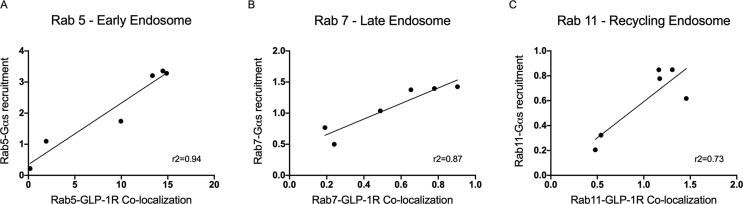
Results: Relative to native and acylated GLP-1 agonists, MAR709 and tirzepatide showed preserved maximal cAMP production despite partial Gαs recruitment paralleled by diminished ligand-induced receptor internalization at both target receptors. Despite MAR709's lower internalization rate, GLP-1R co-localization with Rab11-associated recycling endosomes was not different between MAR709 and GLP-1R specific mono-agonists.
Conclusions: Our data indicated that MAR709 and tirzepatide induce unique spatiotemporal GLP-1 and GIP receptor signaling, trafficking, and recycling dynamics relative to native peptides, semaglutide, and matched mono-agonist controls. These findings support the hypothesis that the structure of GLP-1/GIP dual-agonists confer a biased agonism that, in addition to its influence on intracellular signaling, uniquely modulates receptor trafficking.
Keywords: Biased agonism; Dual-agonists; GIPR; GLP-1R; Receptor Internalization; Receptor Trafficking.
Copyright © 2021 The Authors. Published by Elsevier GmbH.. All rights reserved.
Figures
References
-
- Deacon C.F., Johnsen A.H., Holst J.J. Degradation of glucagon-like peptide-1 by human plasma in vitro yields an N-terminally truncated peptide that is a major endogenous metabolite in vivo. Journal of Clinical Endocrinology & Metabolism. 1995;80:952–957. - PubMed
-
- Kieffer T.J., McIntosh C.H., Pederson R.A. Degradation of glucose-dependent insulinotropic polypeptide and truncated glucagon-like peptide 1 in vitro and in vivo by dipeptidyl peptidase IV. Endocrinology. 1995;136:3585–3596. - PubMed
-
- Mentlein R., Gallwitz B., Schmidt W.E. Dipeptidyl-peptidase IV hydrolyses gastric inhibitory polypeptide, glucagon-like peptide-1 (7-36)amide, peptide histidine methionine and is responsible for their degradation in human serum. European Journal of Biochemistry. 1993;214:829–835. - PubMed
-
- De Meester I., Korom S., Van Damme J., Scharpe S. CD26, let it cut or cut it down. Immunology Today. 1999;20:367–375. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
