

Multisensory interactions regulate feeding behavior in Drosophila
- PMID: 33558226
- PMCID: PMC7896327
- DOI: 10.1073/pnas.2004523118
Multisensory interactions regulate feeding behavior in Drosophila
Abstract
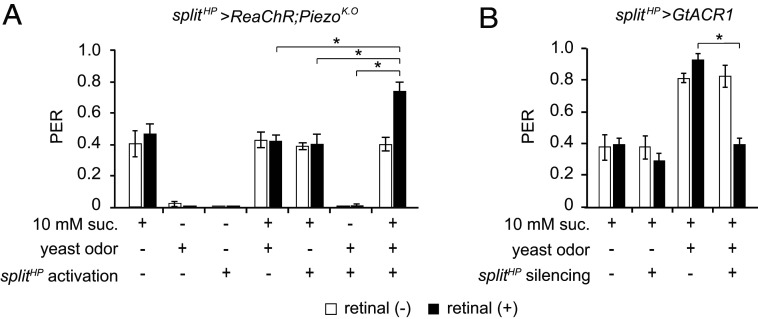
The integration of two or more distinct sensory cues can help animals make more informed decisions about potential food sources, but little is known about how feeding-related multimodal sensory integration happens at the cellular and molecular levels. Here, we show that multimodal sensory integration contributes to a stereotyped feeding behavior in the model organism Drosophila melanogaster Simultaneous olfactory and mechanosensory inputs significantly influence a taste-evoked feeding behavior called the proboscis extension reflex (PER). Olfactory and mechanical information are mediated by antennal Or35a neurons and leg hair plate mechanosensory neurons, respectively. We show that the controlled delivery of three different sensory cues can produce a supra-additive PER via the concurrent stimulation of olfactory, taste, and mechanosensory inputs. We suggest that the fruit fly is a versatile model system to study multisensory integration related to feeding, which also likely exists in vertebrates.
Keywords: Drosophila; mechanosensation; multisensory integration; olfaction; taste.
Conflict of interest statement
The authors declare no competing interest.
Figures
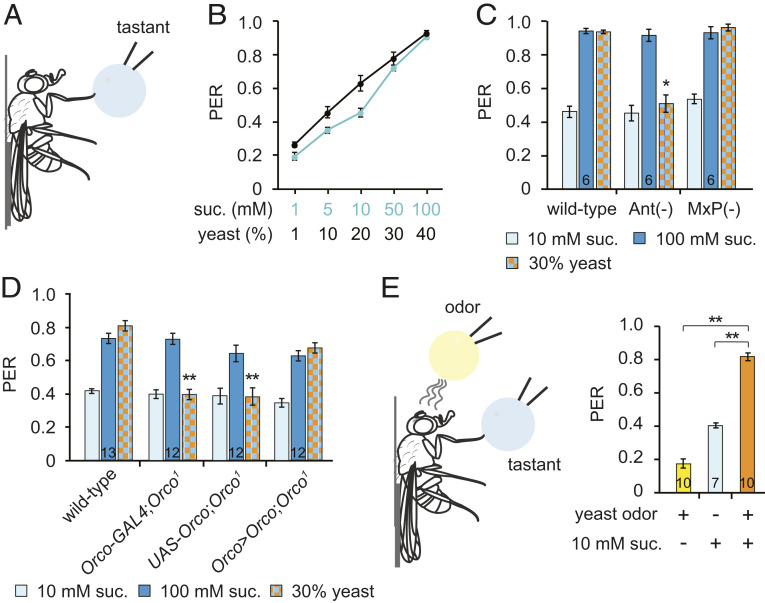
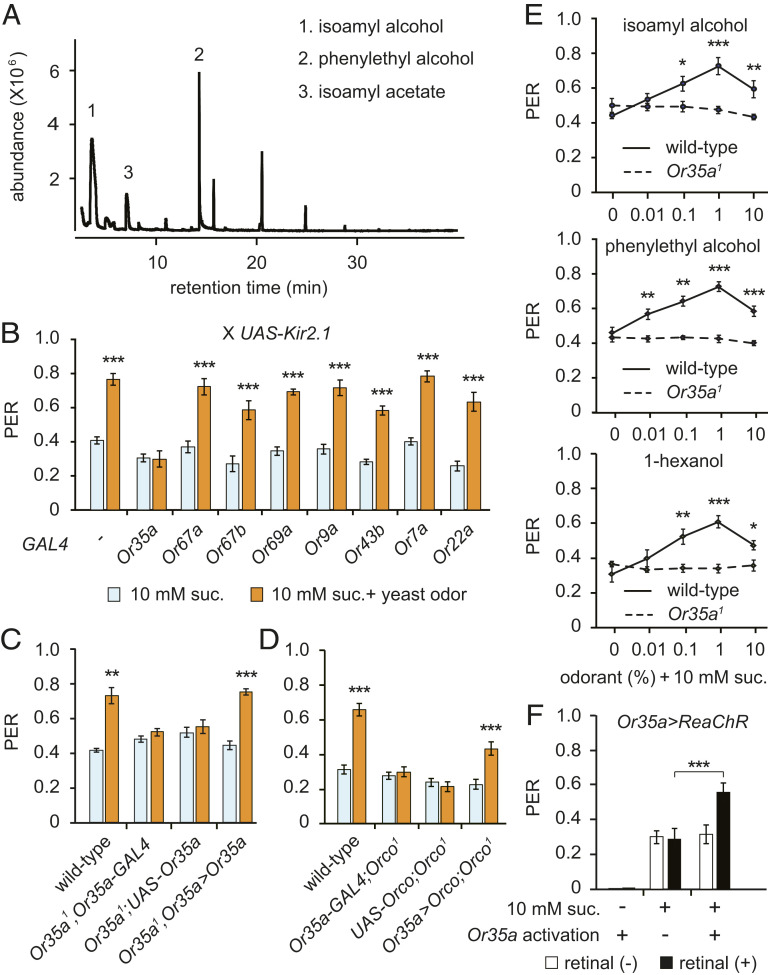
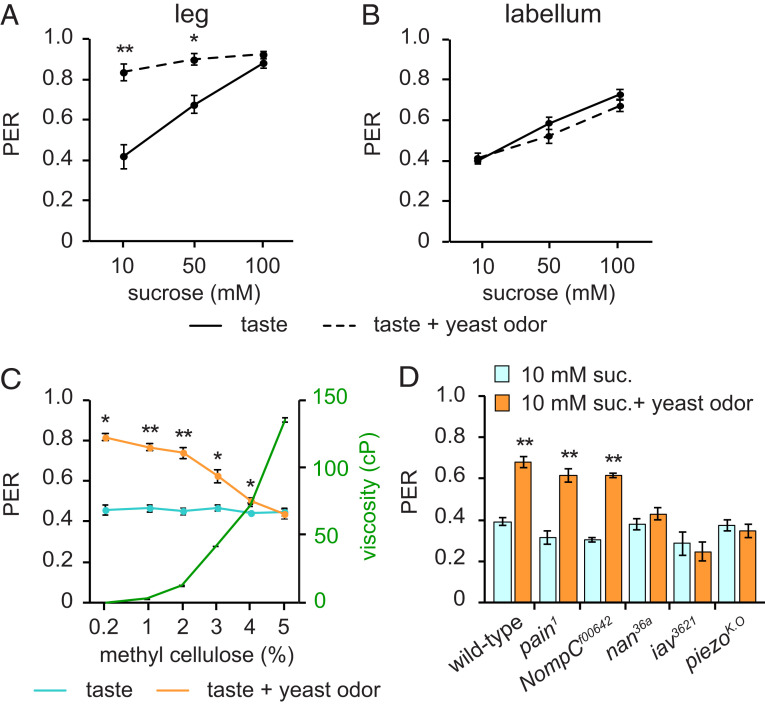
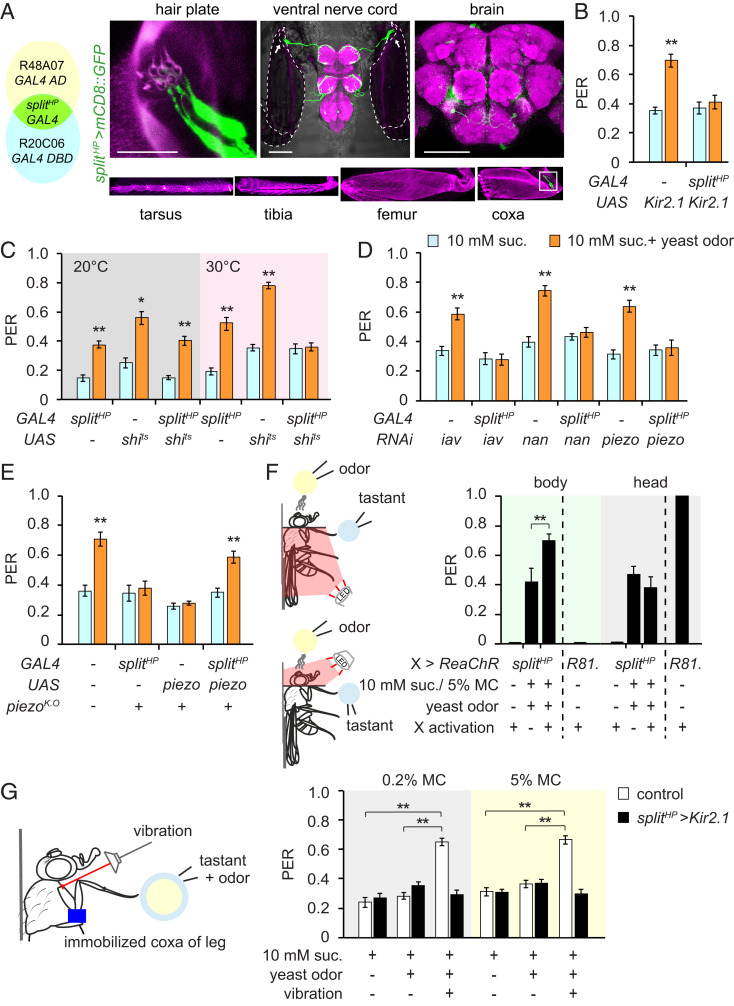
References
-
- Spence C., Multisensory flavor perception. Cell 161, 24–35 (2015). - PubMed
-
- Frank R. A., Byram J., Taste smell interactions are tastant and odorant dependent. Chem. Senses 13, 445–455 (1988).
-
- Paradis S., Sweeney S. T., Davis G. W., Homeostatic control of presynaptic release is triggered by postsynaptic membrane depolarization. Neuron 30, 737–749 (2001). - PubMed
-
- Rozin P., “Taste-smell confusions” and the duality of the olfactory sense. Percept. Psychophys. 31, 397–401 (1982). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
