

Ubiquitin-Specific Protease 6 Functions as a Tumor Suppressor in Ewing Sarcoma through Immune Activation
- PMID: 33558334
- PMCID: PMC8137534
- DOI: 10.1158/0008-5472.CAN-20-1458
Ubiquitin-Specific Protease 6 Functions as a Tumor Suppressor in Ewing Sarcoma through Immune Activation
Abstract
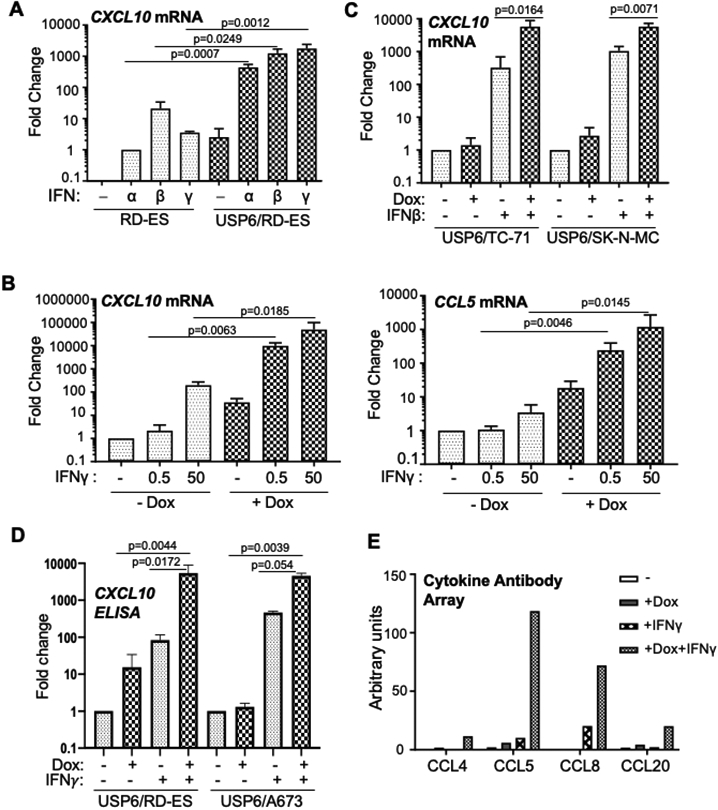
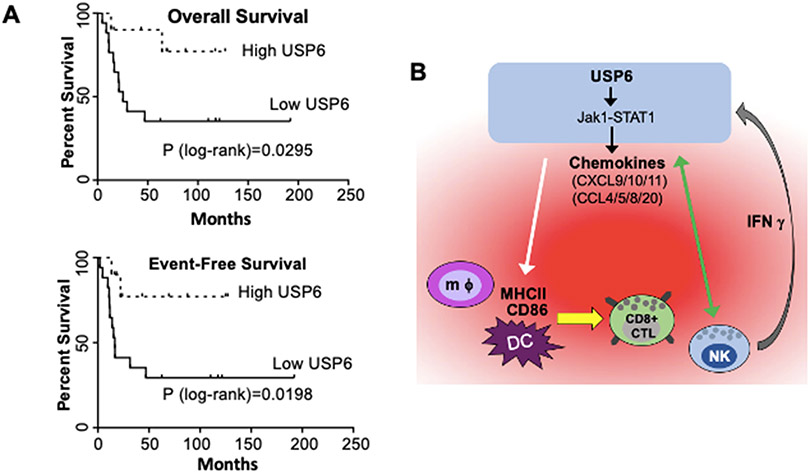
Ewing sarcoma is the second most common pediatric bone cancer, with a 5-year survival rate for metastatic disease of only 20%. Recent work indicates that survival is strongly correlated with high levels of tumor-infiltrating lymphocytes (TIL), whose abundance is associated with IFN-inducible chemokines CXCL10 and CCL5. However, the tumor-intrinsic factors that drive chemokine production and TIL recruitment have not been fully elucidated. We previously showed that ubiquitin-specific protease 6 (USP6) directly deubiquitinates and stabilizes Jak1, thereby inducing an IFN signature in Ewing sarcoma cells. Here, we show that this gene set comprises chemokines associated with immunostimulatory, antitumorigenic functions, including CXCL10 and CCL5. USP6 synergistically enhanced chemokine production in response to exogenous IFN by inducing surface upregulation of IFNAR1 and IFNGR1. USP6-expressing Ewing sarcoma cells stimulated migration of primary human monocytes and T lymphocytes and triggered activation of natural killer (NK) cells in vitro. USP6 inhibited Ewing sarcoma xenograft growth in nude but not NSG mice and was accompanied by increased intratumoral chemokine production and infiltration and activation of NK cells, dendritic cells, and macrophages, consistent with a requirement for innate immune cells in mediating the antitumorigenic effects of USP6. High USP6 expression in patients with Ewing sarcoma was associated with chemokine production, immune infiltration, and improved survival. This work reveals a previously unrecognized tumor-suppressive function for USP6, which engenders an immunostimulatory microenvironment through pleiotropic effects on multiple immune lineages. This further raises the possibility that USP6 activity may be harnessed to create a "hot" tumor microenvironment in immunotherapy. SIGNIFICANCE: This study reveals a novel tumor-suppressive function for USP6 by inducing an immunostimulatory microenvironment, suggesting that USP6 activity may be exploited to enhance immunotherapy regimens.
©2021 American Association for Cancer Research.
Conflict of interest statement
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
