

Darwinian properties and their trade-offs in autocatalytic RNA reaction networks
- PMID: 33558542
- PMCID: PMC7870898
- DOI: 10.1038/s41467-021-21000-1
Darwinian properties and their trade-offs in autocatalytic RNA reaction networks
Abstract
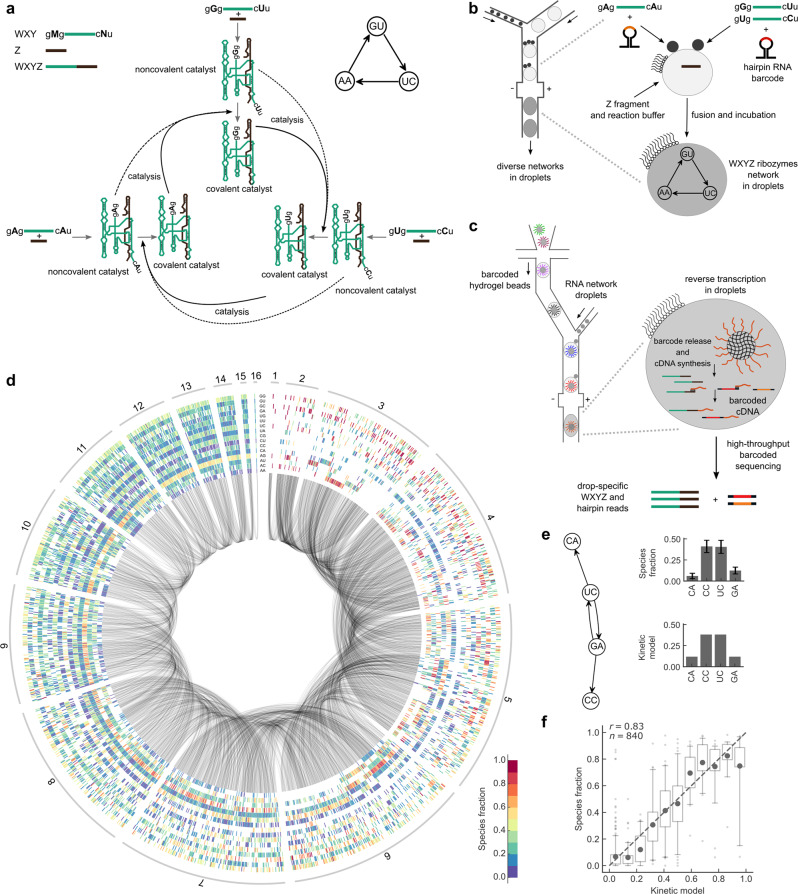
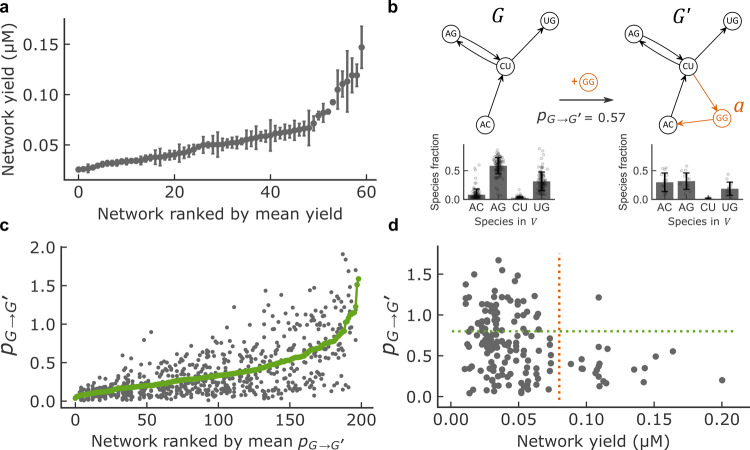
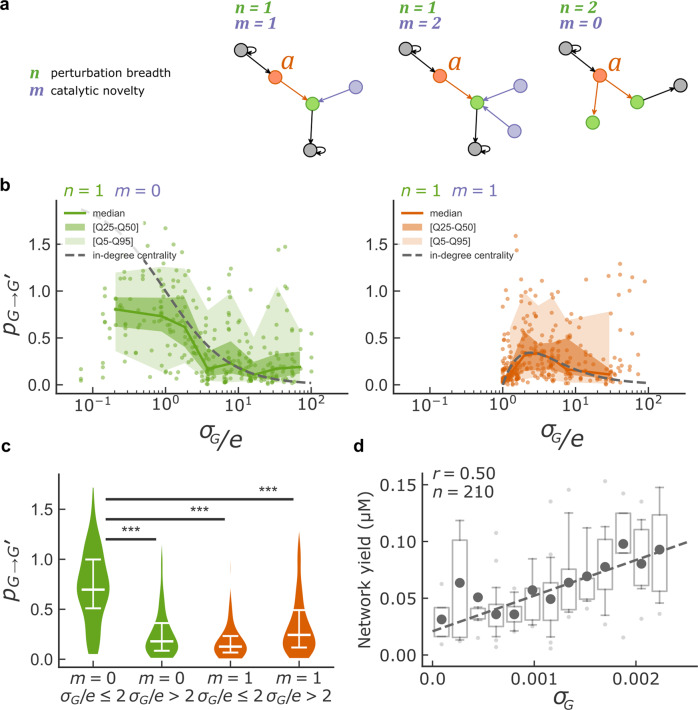
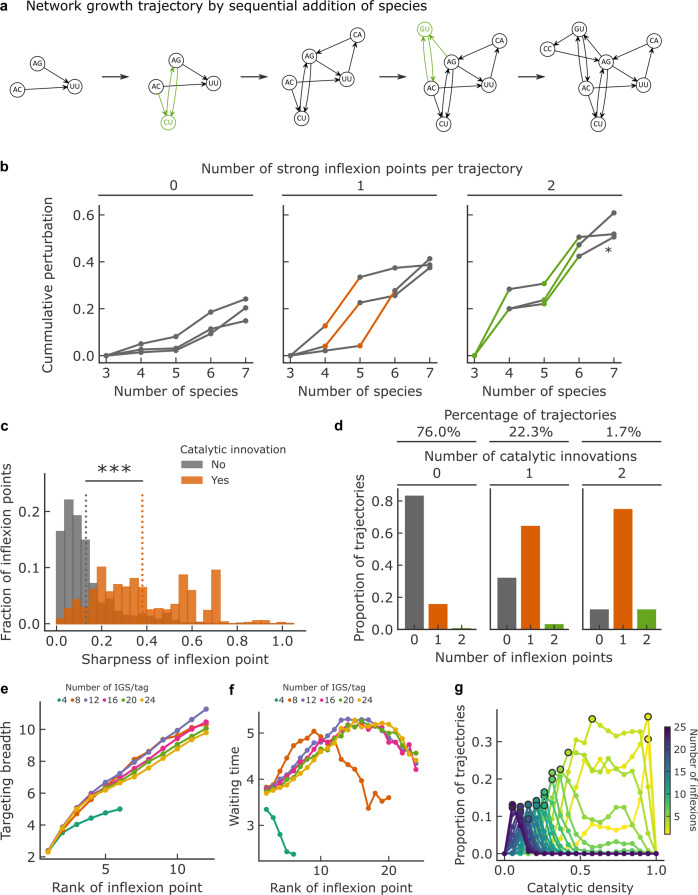
Discovering autocatalytic chemistries that can evolve is a major goal in systems chemistry and a critical step towards understanding the origin of life. Autocatalytic networks have been discovered in various chemistries, but we lack a general understanding of how network topology controls the Darwinian properties of variation, differential reproduction, and heredity, which are mediated by the chemical composition. Using barcoded sequencing and droplet microfluidics, we establish a landscape of thousands of networks of RNAs that catalyze their own formation from fragments, and derive relationships between network topology and chemical composition. We find that strong variations arise from catalytic innovations perturbing weakly connected networks, and that growth increases with global connectivity. These rules imply trade-offs between reproduction and variation, and between compositional persistence and variation along trajectories of network complexification. Overall, connectivity in reaction networks provides a lever to balance variation (to explore chemical states) with reproduction and heredity (persistence being necessary for selection to act), as required for chemical evolution.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Butlerow A. Formation synthétique d’une substance sucrée. Comp. Rend. Acad. Sci. 1861;53:145–147.
Publication types
MeSH terms
Substances
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
