

Fully defined human pluripotent stem cell-derived microglia and tri-culture system model C3 production in Alzheimer's disease
- PMID: 33558694
- PMCID: PMC8382543
- DOI: 10.1038/s41593-020-00796-z
Fully defined human pluripotent stem cell-derived microglia and tri-culture system model C3 production in Alzheimer's disease
Abstract
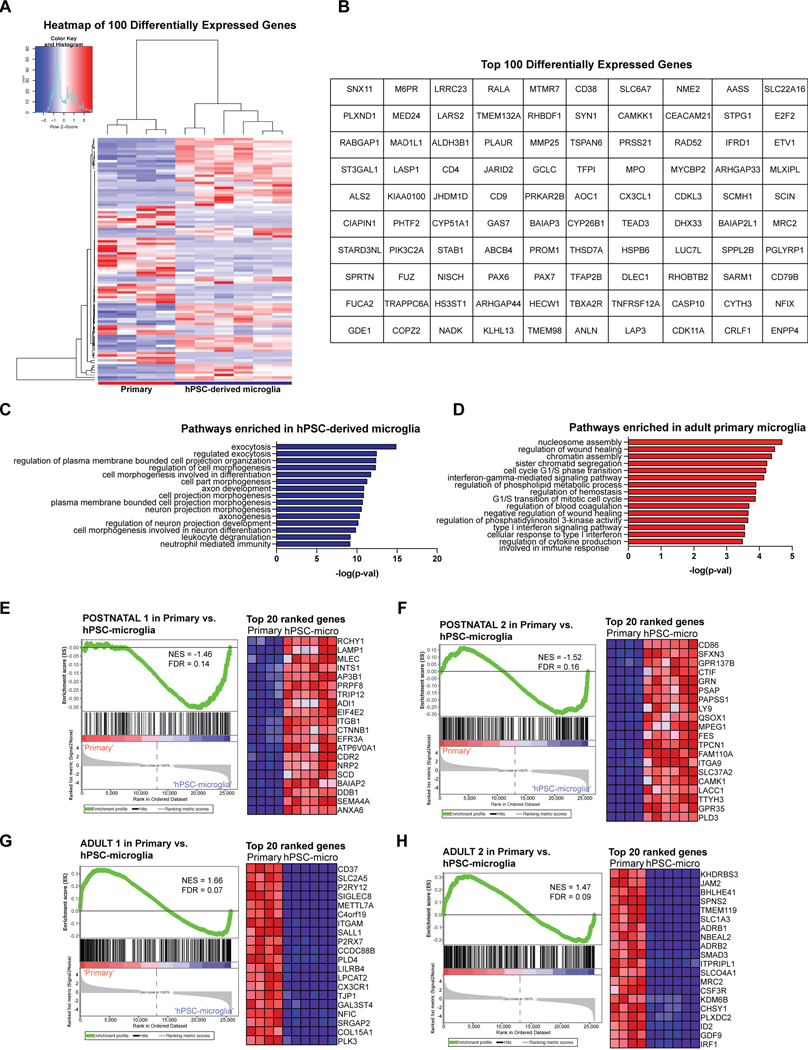
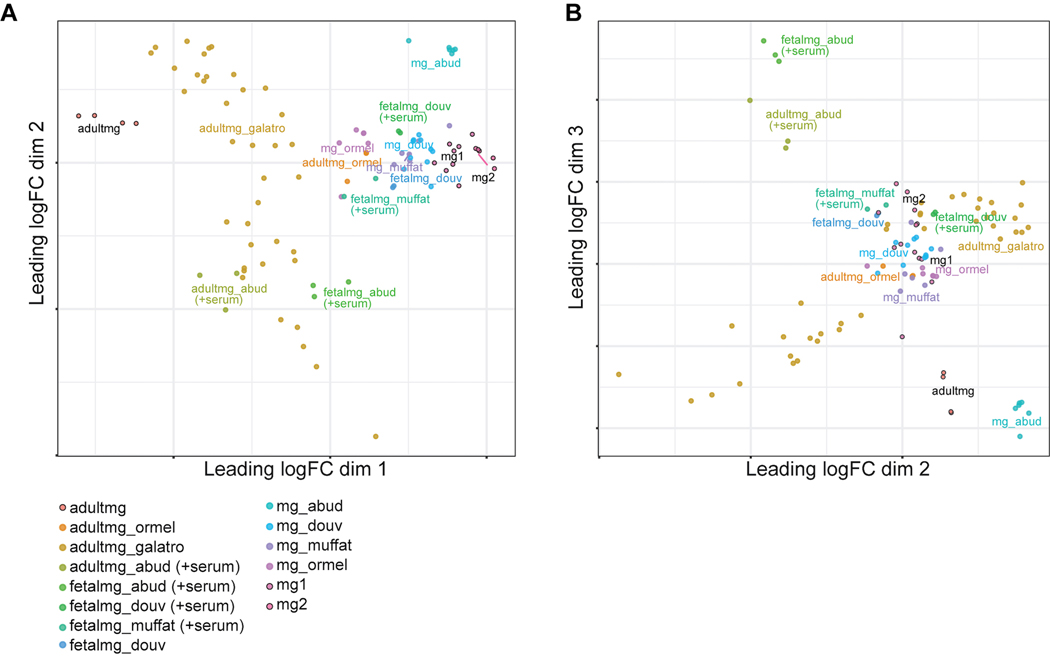
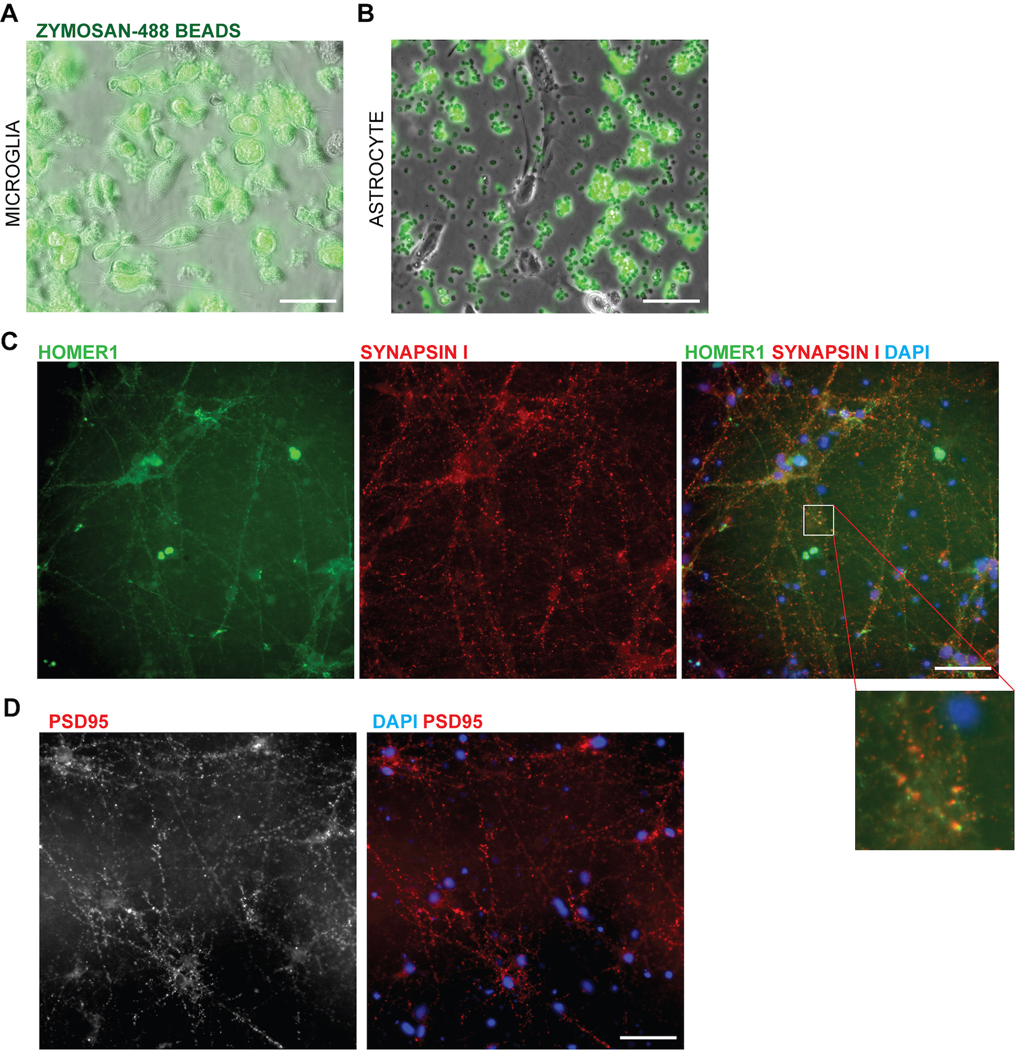
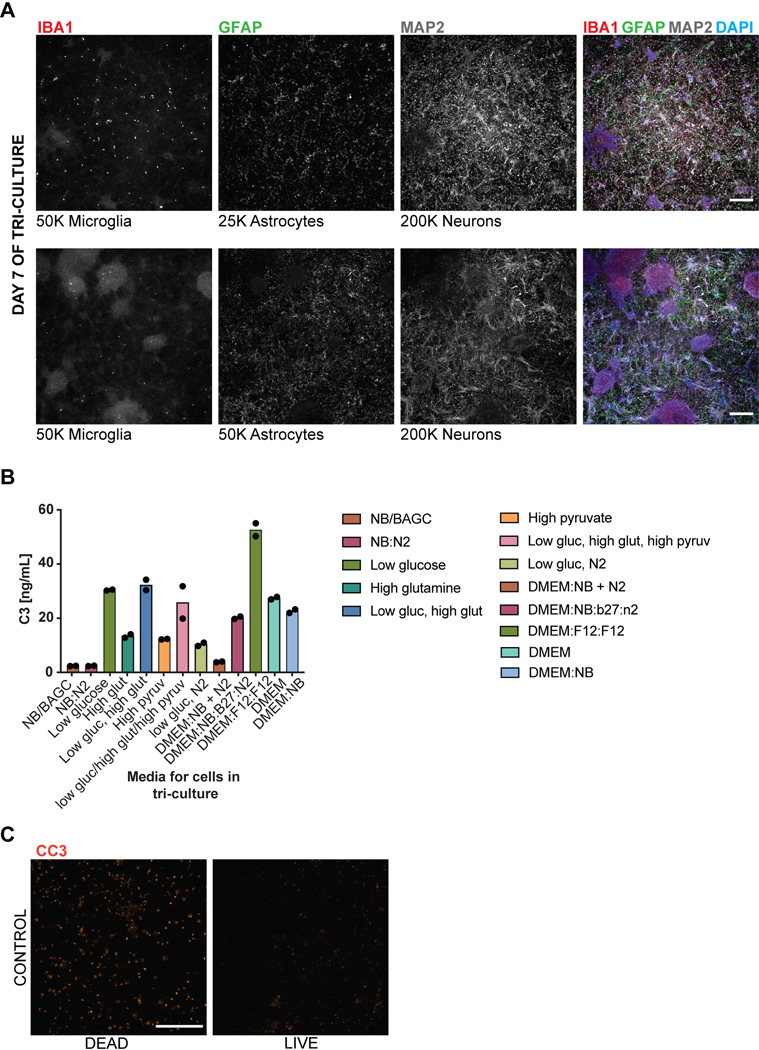
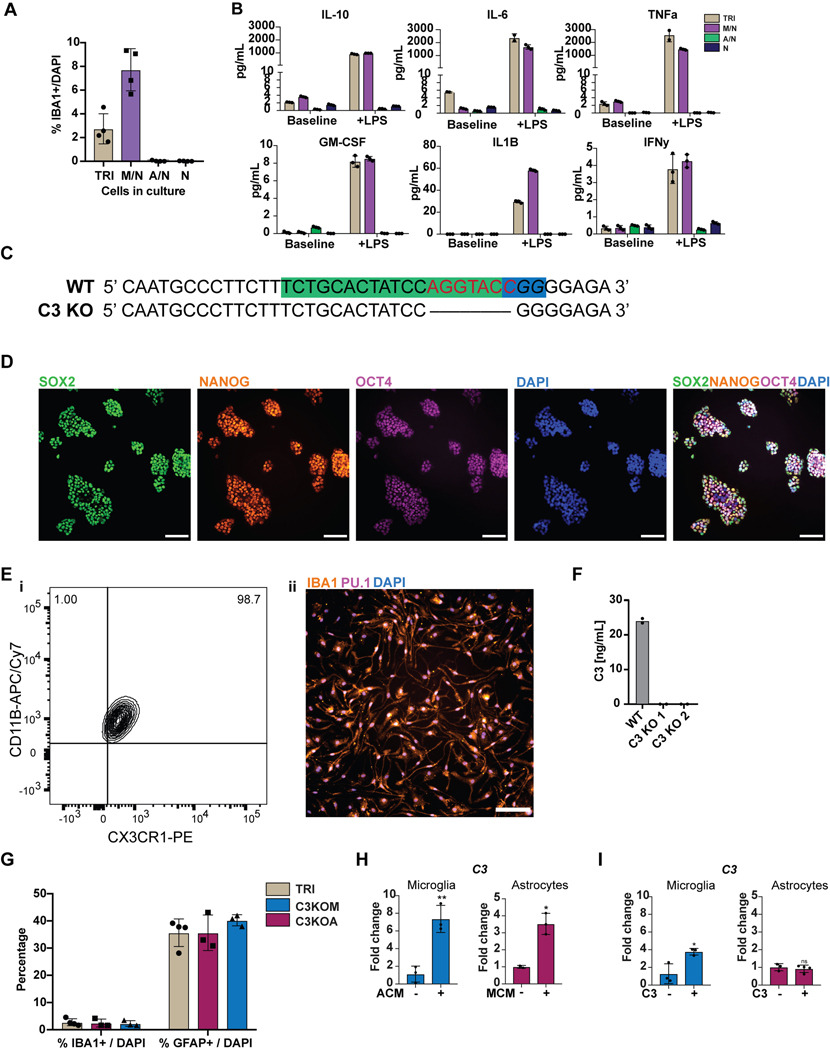
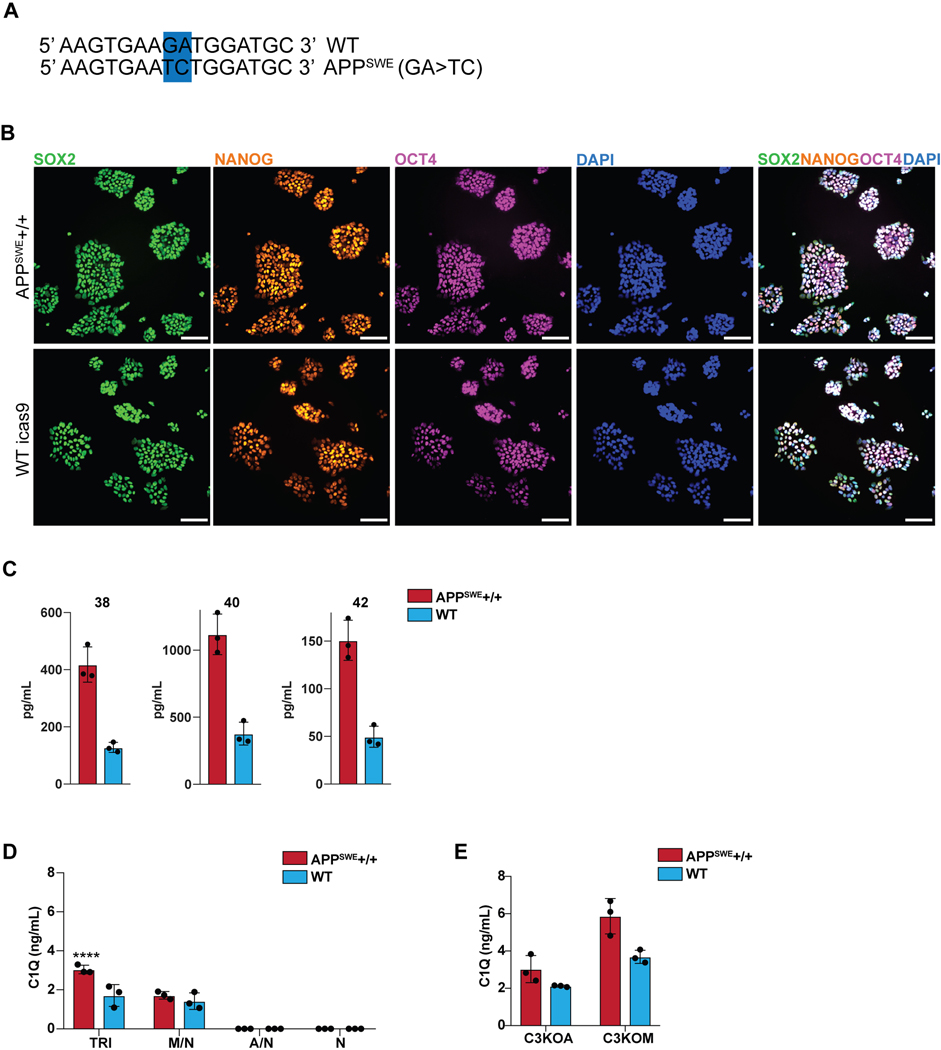
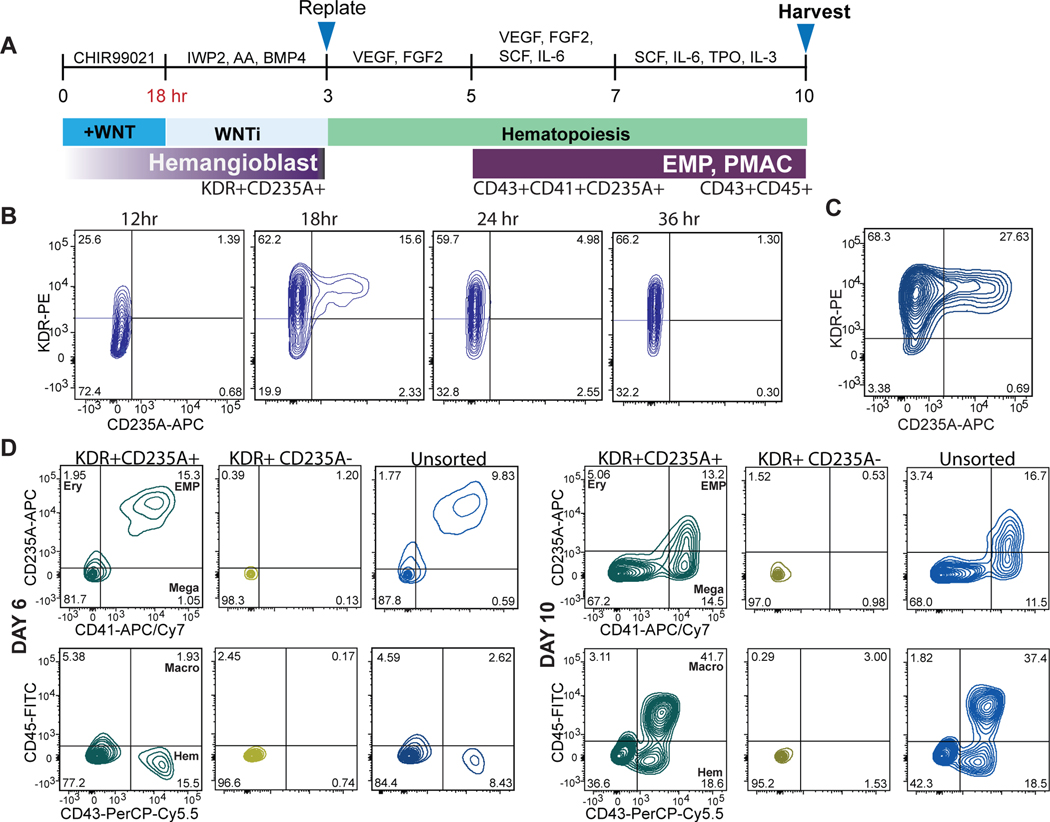
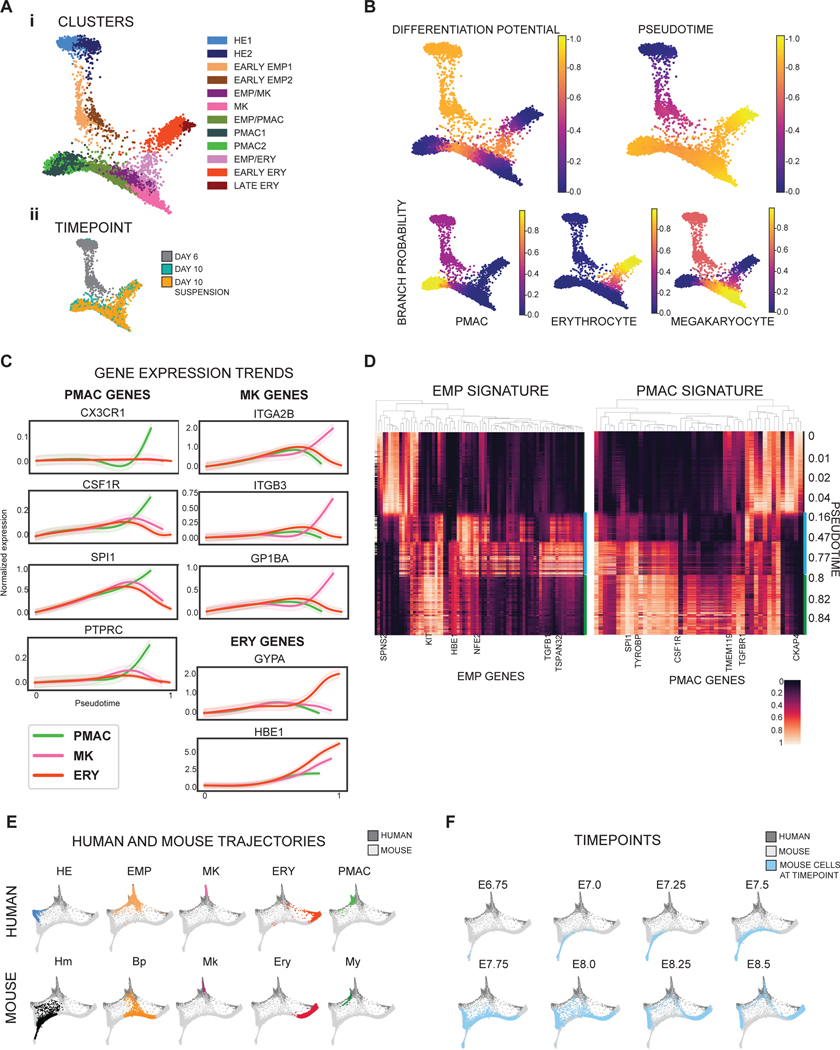
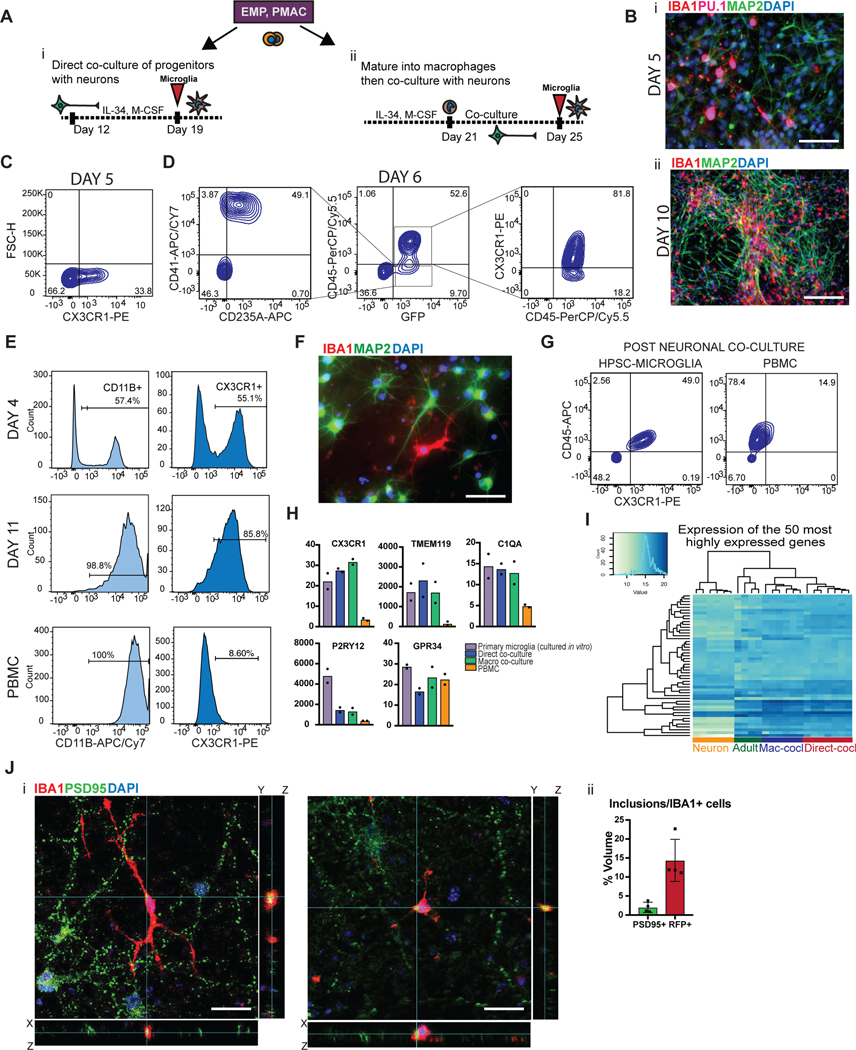
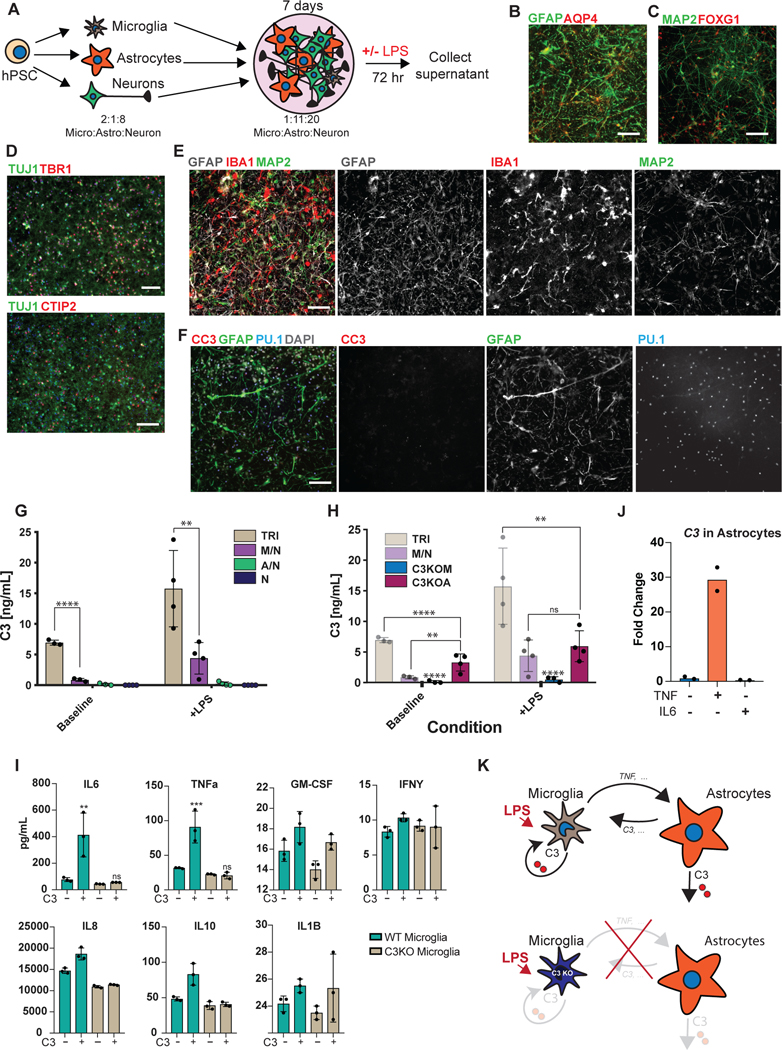
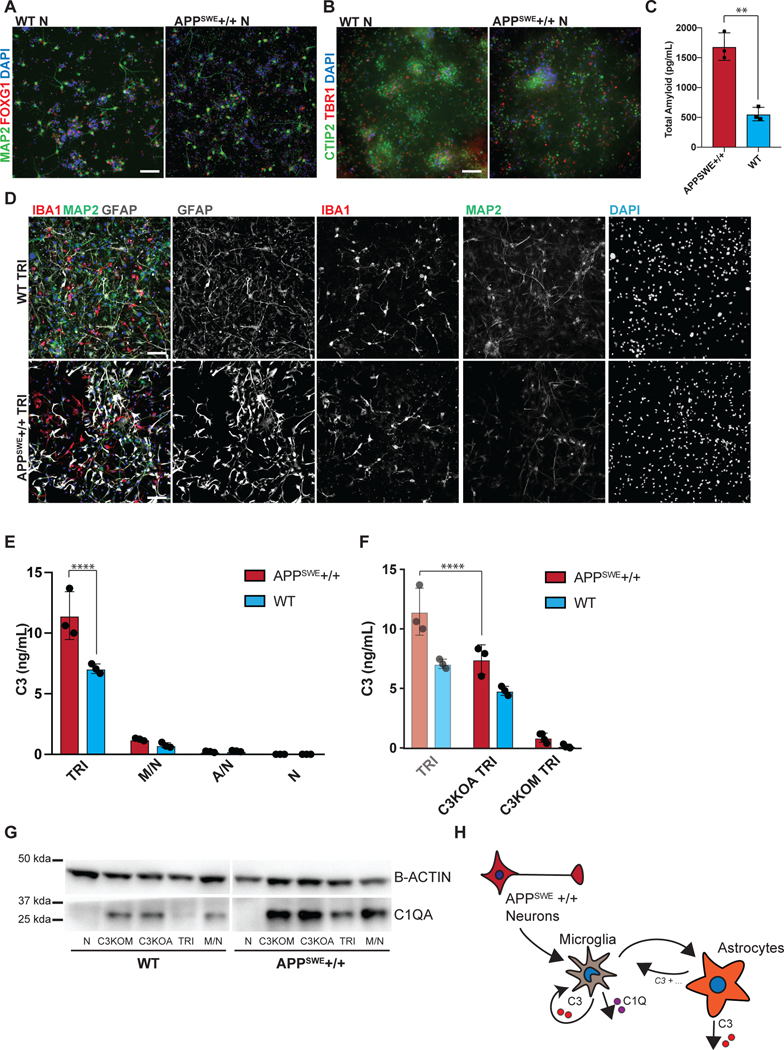
Aberrant inflammation in the CNS has been implicated as a major player in the pathogenesis of human neurodegenerative disease. We developed a new approach to derive microglia from human pluripotent stem cells (hPSCs) and built a defined hPSC-derived tri-culture system containing pure populations of hPSC-derived microglia, astrocytes, and neurons to dissect cellular cross-talk along the neuroinflammatory axis in vitro. We used the tri-culture system to model neuroinflammation in Alzheimer's disease with hPSCs harboring the APPSWE+/+ mutation and their isogenic control. We found that complement C3, a protein that is increased under inflammatory conditions and implicated in synaptic loss, is potentiated in tri-culture and further enhanced in APPSWE+/+ tri-cultures due to microglia initiating reciprocal signaling with astrocytes to produce excess C3. Our study defines the major cellular players contributing to increased C3 in Alzheimer's disease and presents a broadly applicable platform to study neuroinflammation in human disease.
Figures
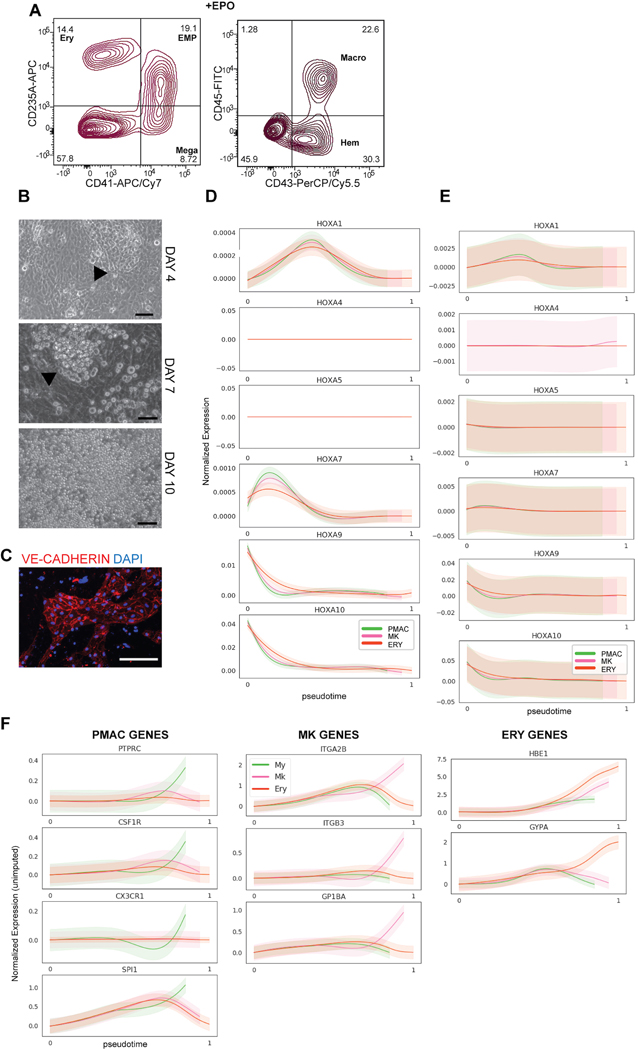
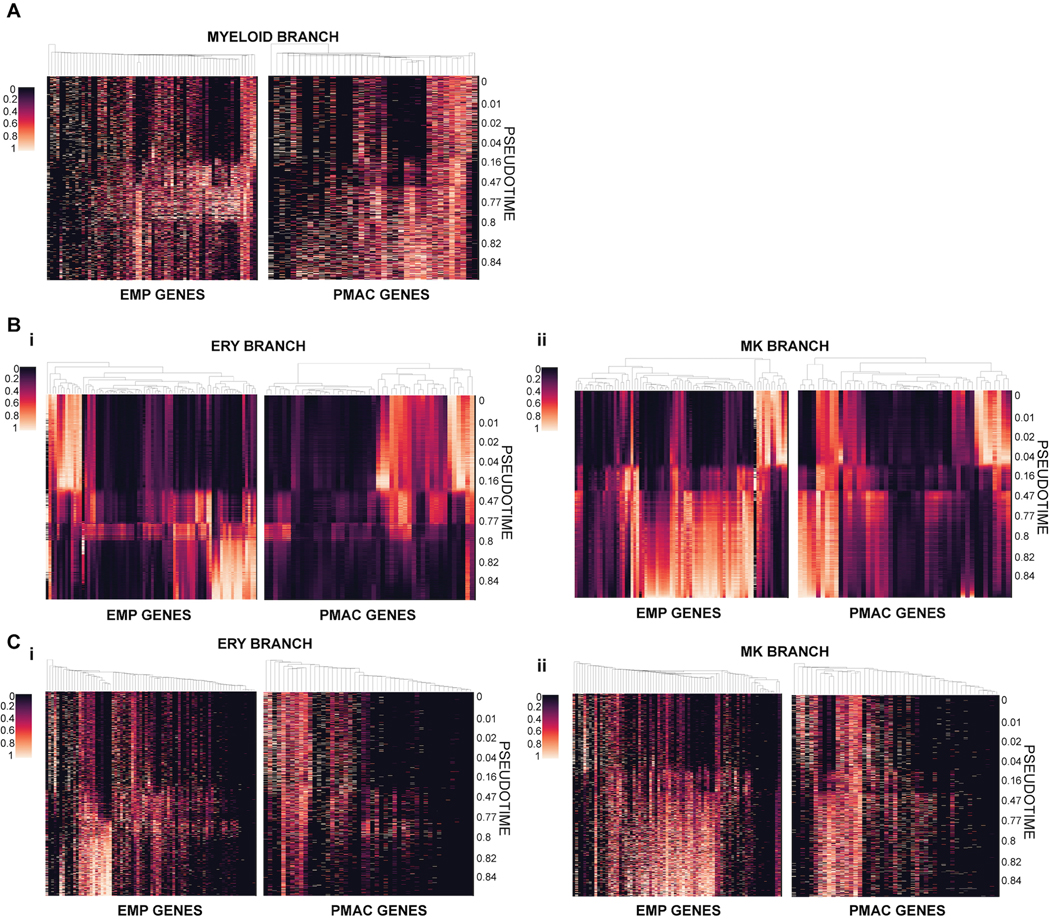
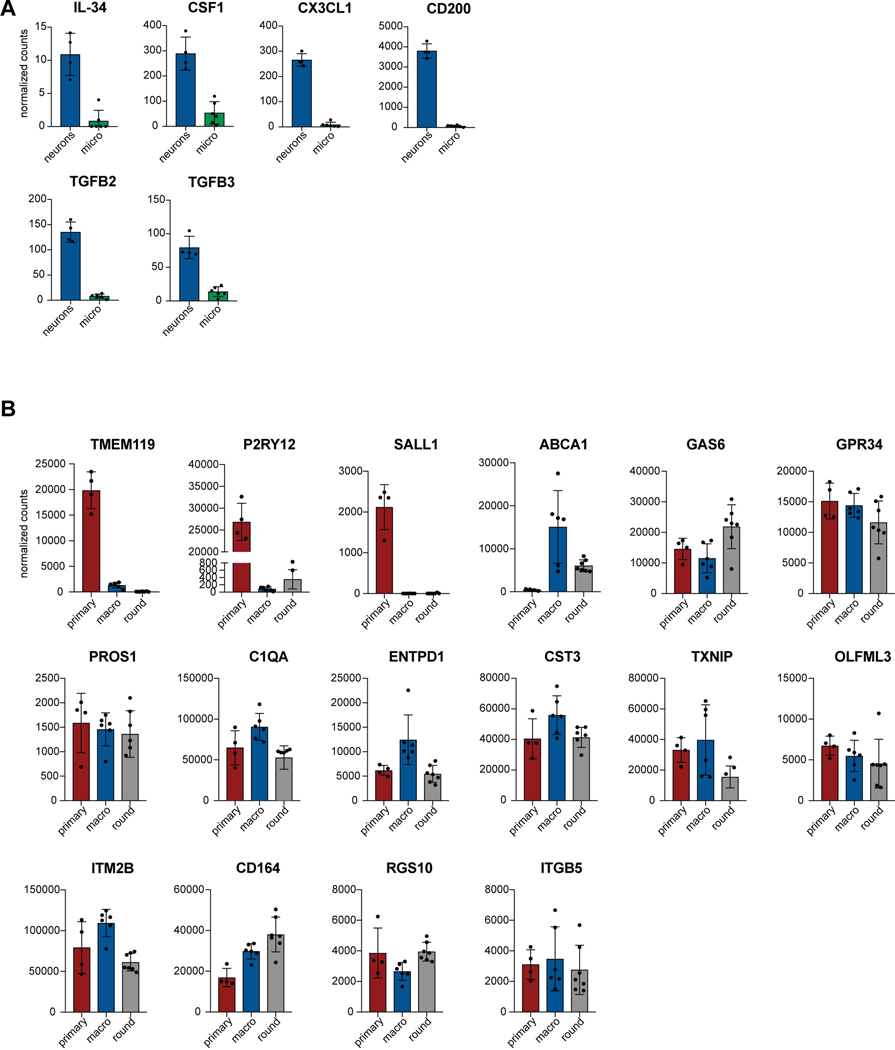
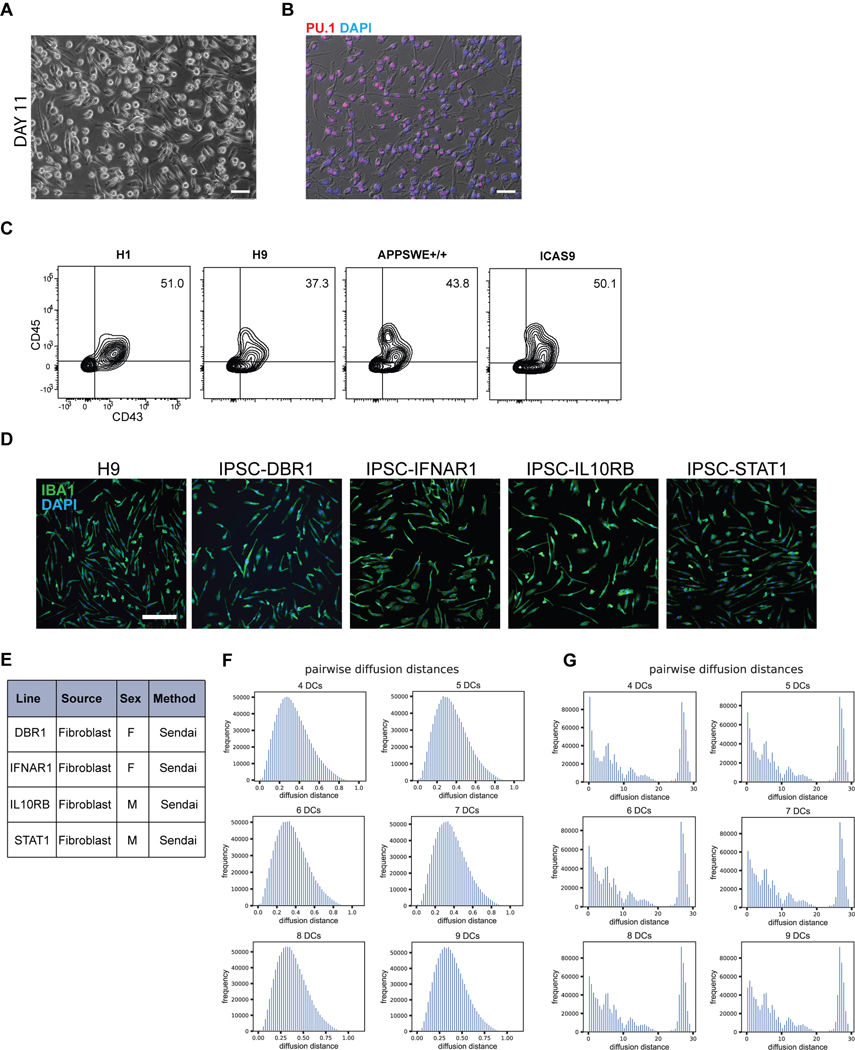
References
CITED REFERENCES
METHODS REFERENCES
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
