

αβγδ T cells play a vital role in fetal human skin development and immunity
- PMID: 33561194
- PMCID: PMC7876551
- DOI: 10.1084/jem.20201189
αβγδ T cells play a vital role in fetal human skin development and immunity
Abstract
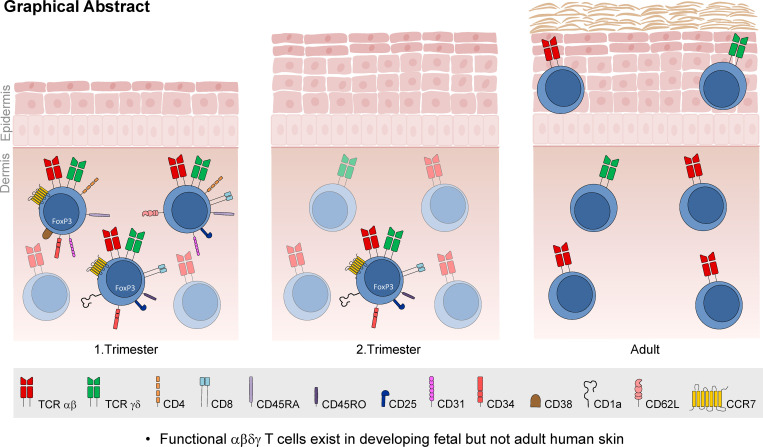
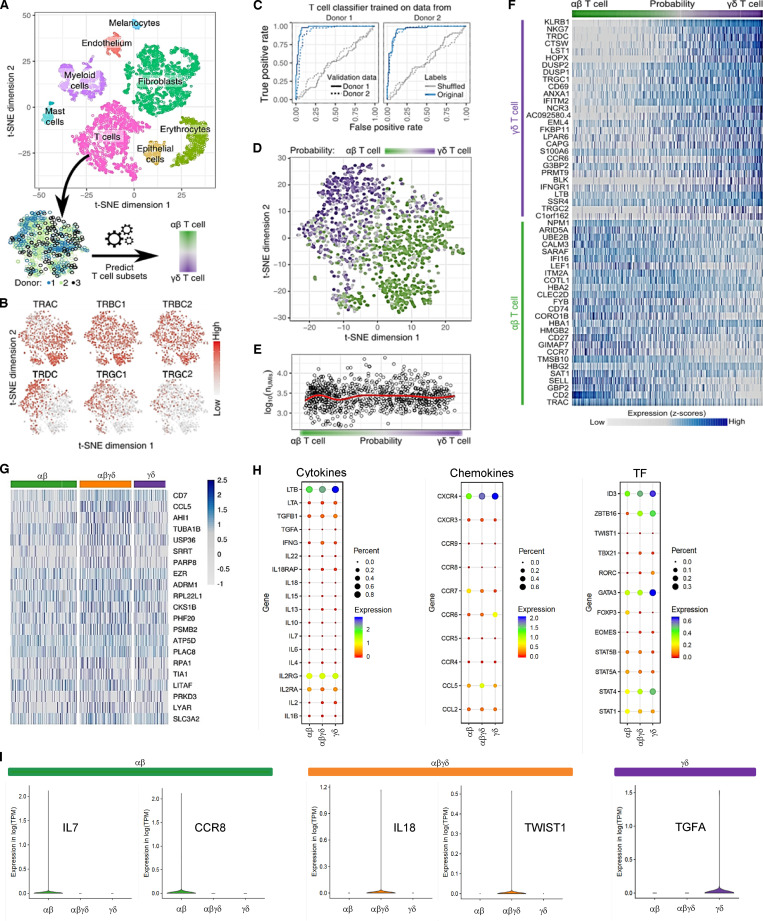
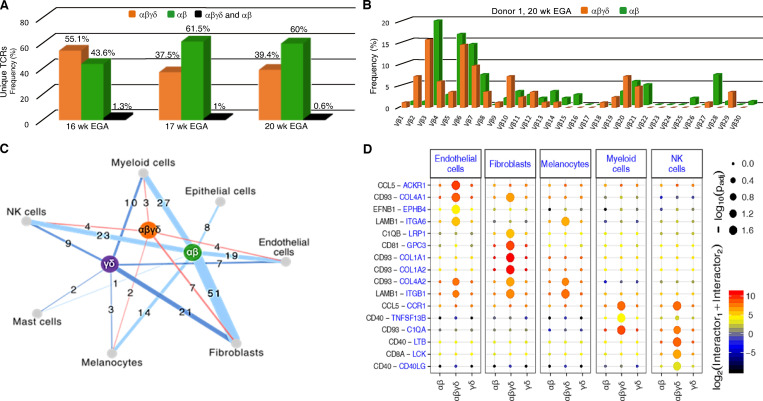
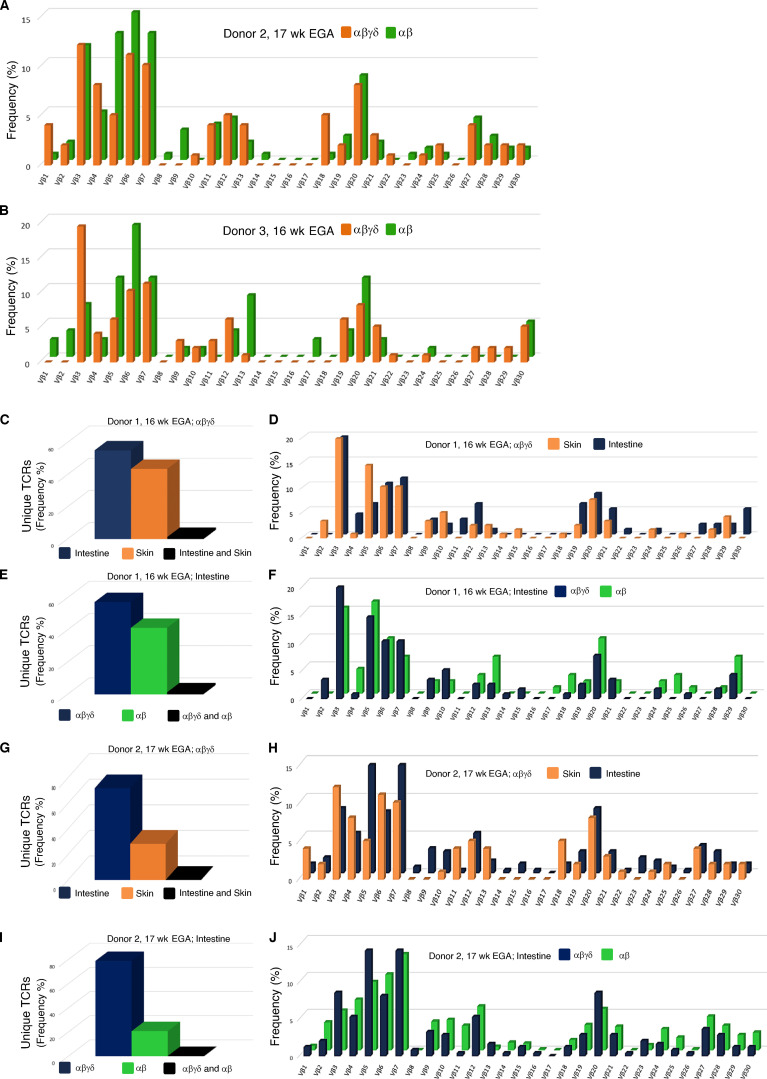
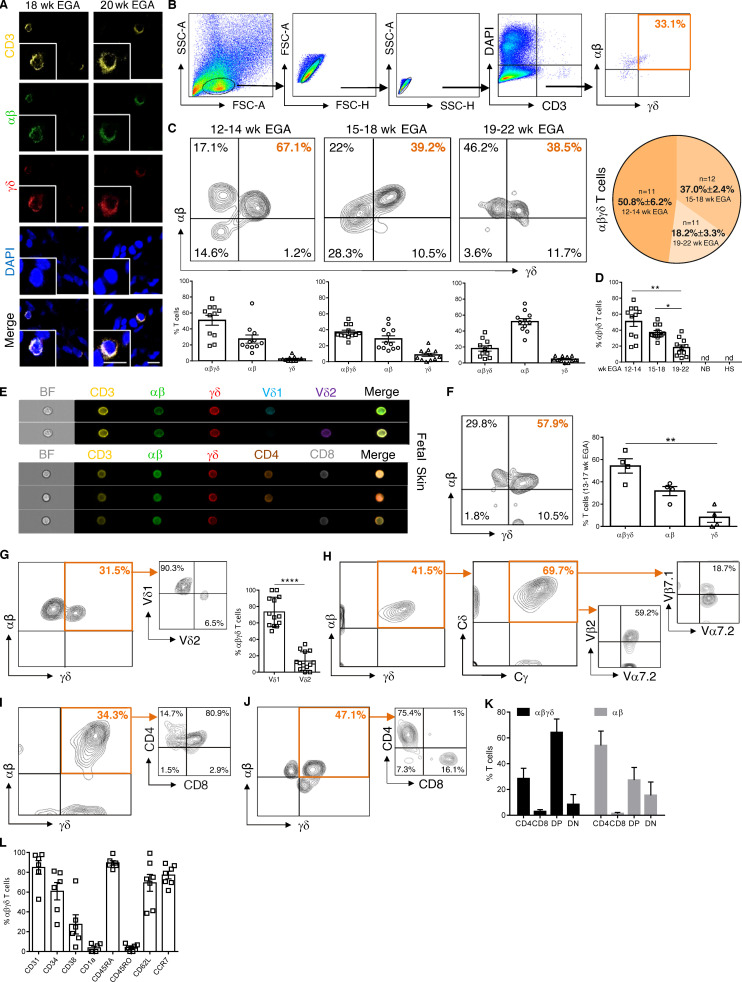
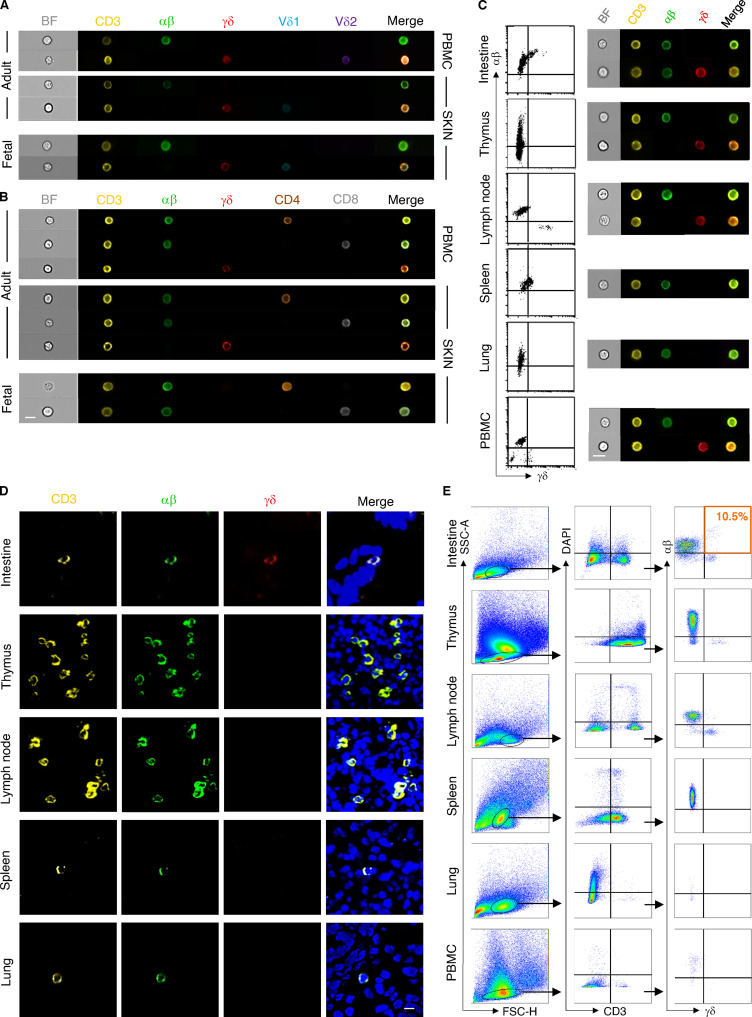
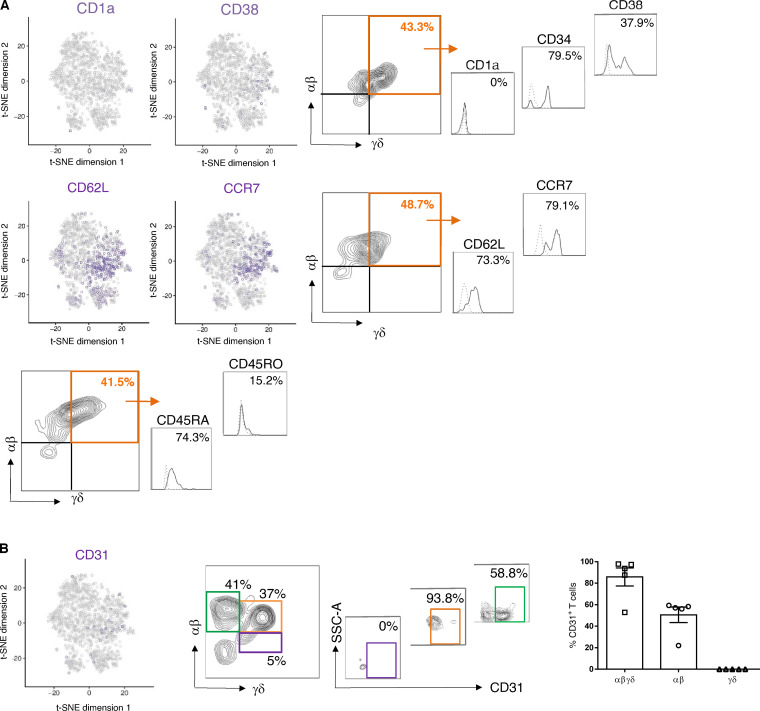
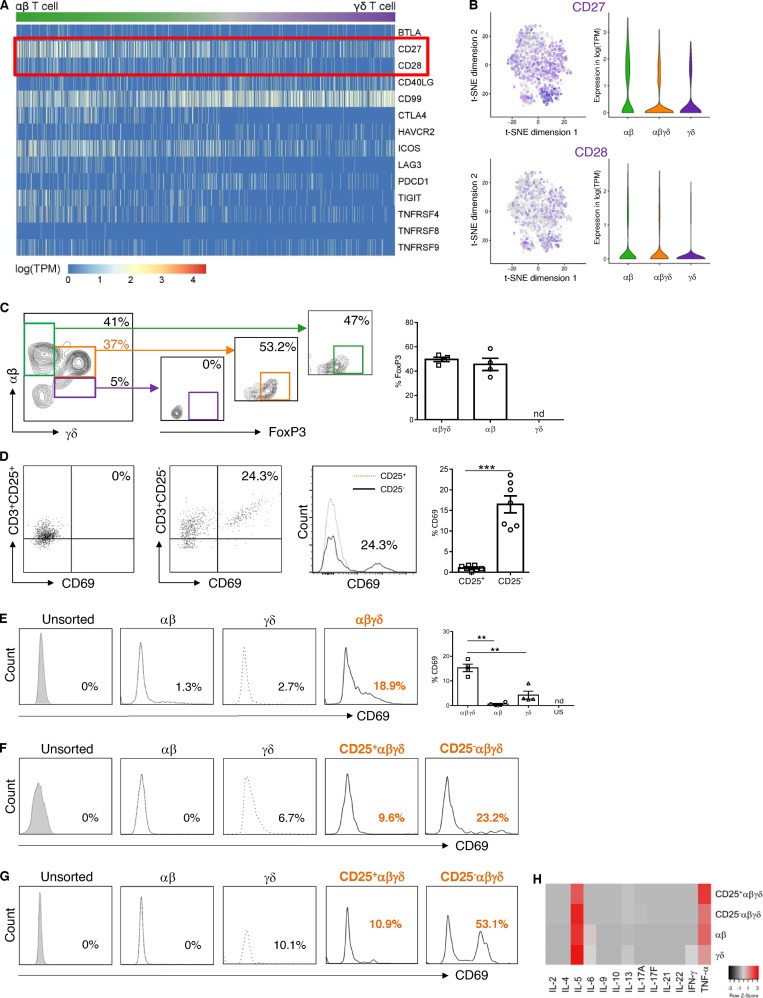
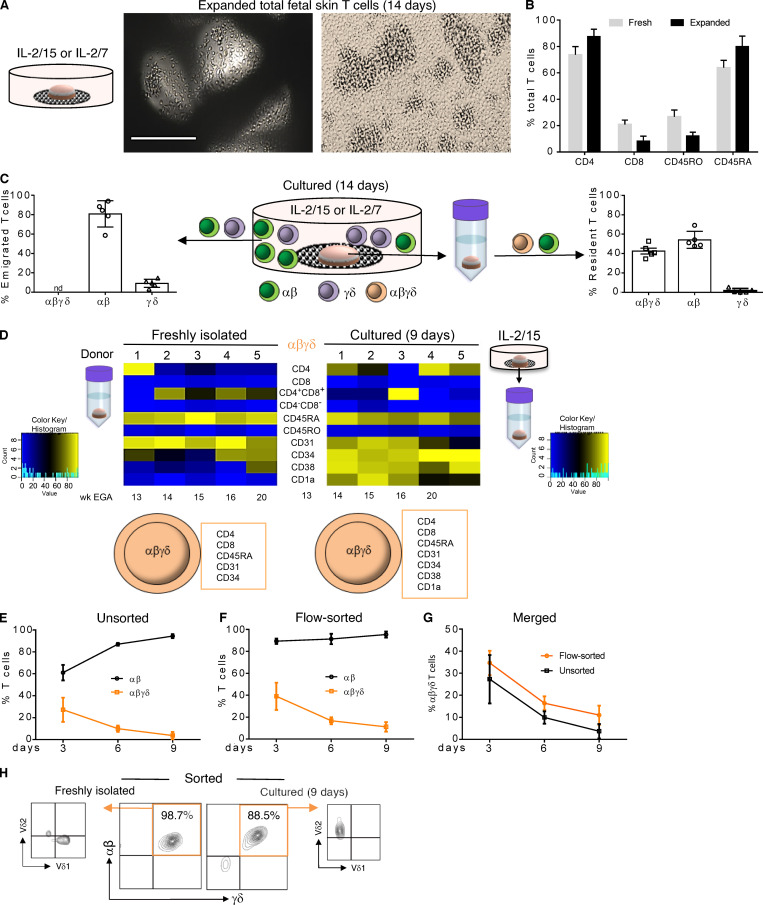
T cells in human skin play an important role in the immune defense against pathogens and tumors. T cells are present already in fetal skin, where little is known about their cellular phenotype and biological function. Using single-cell analyses, we identified a naive T cell population expressing αβ and γδ T cell receptors (TCRs) that was enriched in fetal skin and intestine but not detected in other fetal organs and peripheral blood. TCR sequencing data revealed that double-positive (DP) αβγδ T cells displayed little overlap of CDR3 sequences with single-positive αβ T cells. Gene signatures, cytokine profiles and in silico receptor-ligand interaction studies indicate their contribution to early skin development. DP αβγδ T cells were phosphoantigen responsive, suggesting their participation in the protection of the fetus against pathogens in intrauterine infections. Together, our analyses unveil a unique cutaneous T cell type within the native skin microenvironment and point to fundamental differences in the immune surveillance between fetal and adult human skin.
© 2021 Reitermaier et al.
Conflict of interest statement
Disclosures: The authors declare no competing interests exist.
Figures
References
-
- Byrne, J.A., Stankovic A.K., and Cooper M.D.. 1994. A novel subpopulation of primed T cells in the human fetus. J. Immunol. 152:3098–3106. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
