

Motility of efferent duct cilia aids passage of sperm cells through the male reproductive system
- PMID: 33561200
- PMCID: PMC7936721
- DOI: 10.1093/molehr/gaab009
Motility of efferent duct cilia aids passage of sperm cells through the male reproductive system
Abstract
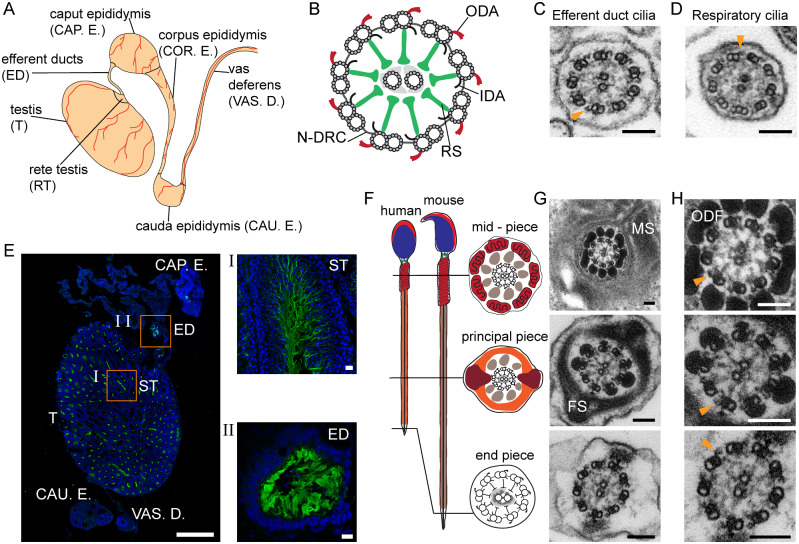
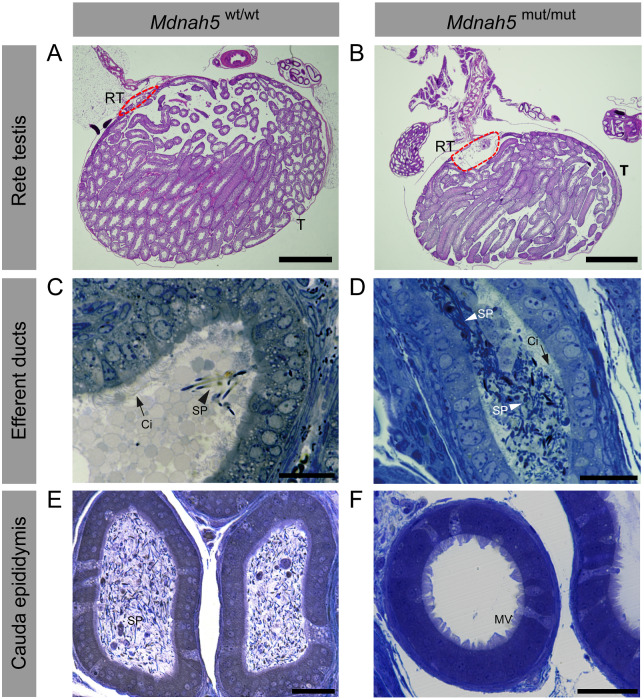
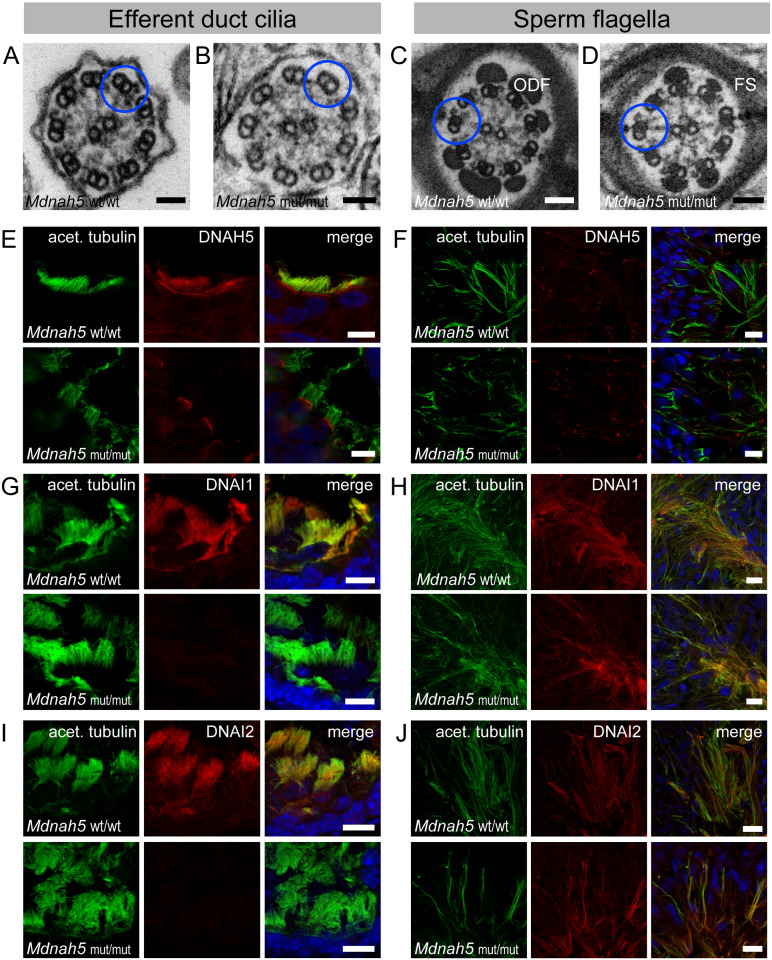
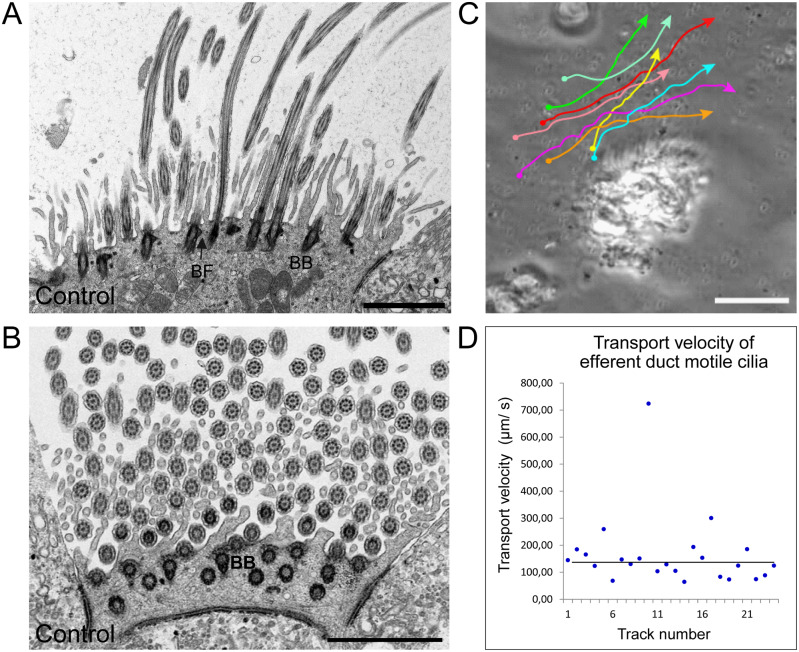
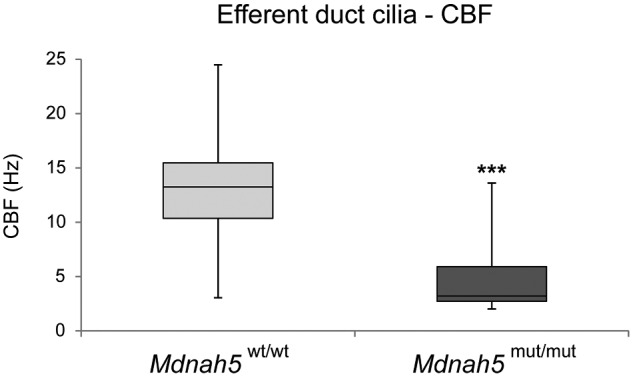
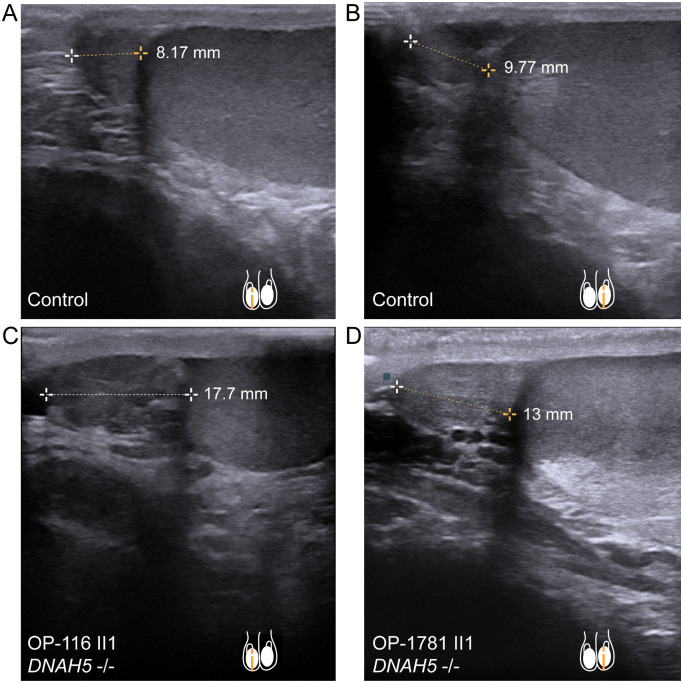
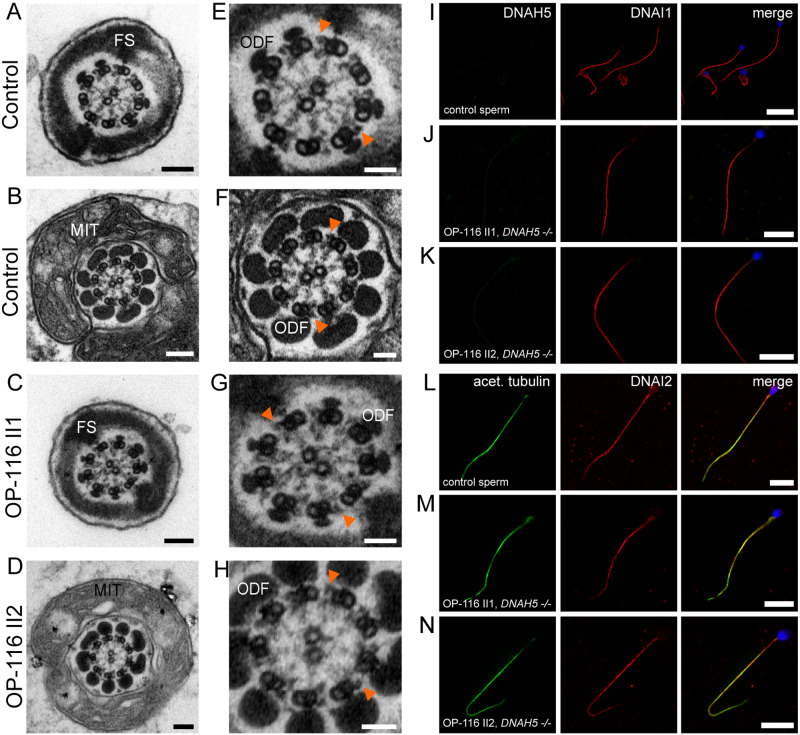
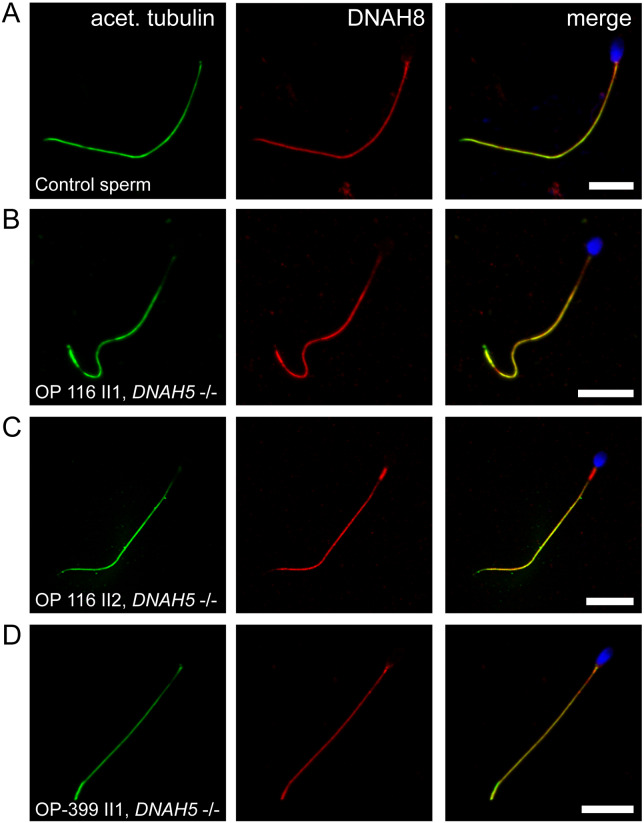
Motile cilia line the efferent ducts of the mammalian male reproductive tract. Several recent mouse studies have demonstrated that a reduced generation of multiple motile cilia in efferent ducts is associated with obstructive oligozoospermia and fertility issues. However, the sole impact of efferent duct cilia dysmotility on male infertility has not been studied so far either in mice or human. Using video microscopy, histological- and ultrastructural analyses, we examined male reproductive tracts of mice deficient for the axonemal motor protein DNAH5: this defect exclusively disrupts the outer dynein arm (ODA) composition of motile cilia but not the ODA composition and motility of sperm flagella. These mice have immotile efferent duct cilia that lack ODAs, which are essential for ciliary beat generation. Furthermore, they show accumulation of sperm in the efferent duct. Notably, the ultrastructure and motility of sperm from these males are unaffected. Likewise, human individuals with loss-of-function DNAH5 mutations present with reduced sperm count in the ejaculate (oligozoospermia) and dilatations of the epididymal head but normal sperm motility, similar to DNAH5 deficient mice. The findings of this translational study demonstrate, in both mice and men, that efferent duct ciliary motility is important for male reproductive fitness and uncovers a novel pathomechanism distinct from primary defects of sperm motility (asthenozoospermia). If future work can identify environmental factors or defects in genes other than DNAH5 that cause efferent duct cilia dysmotility, this will help unravel other causes of oligozoospermia and may influence future practices in genetic and fertility counseling as well as ART.
Keywords: efferent ducts / cilia / sperm / motor proteins / outer dynein arm-defects / DNAH5 / 45 male infertility / oligozoospermia / motile ciliopathy / primary ciliary dyskinesia.
© The Author(s) 2021. Published by Oxford University Press on behalf of European Society of Human Reproduction and Embryology.
Figures
References
-
- Becker O. Ueber Flimmerepithelium im Nebenhoden des Menschen. Wiener Medizinische Wochenschrift 1856;6:184.
-
- Björndahl L, Barratt CLR, Mortimer D, Jouannet P. ‘How to count sperm properly’: checklist for acceptability of studies based on human semen analysis. Hum Reprod 2016;31:227–232. - PubMed
-
- Bracke A, Peeters K, Punjabi U, Hoogewijs D, Dewilde S. A search for molecular mechanisms underlying male idiopathic infertility. Reprod Biomed Online 2018;36:327–339. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
