

The actin networks of chytrid fungi reveal evolutionary loss of cytoskeletal complexity in the fungal kingdom
- PMID: 33561386
- PMCID: PMC8812817
- DOI: 10.1016/j.cub.2021.01.001
The actin networks of chytrid fungi reveal evolutionary loss of cytoskeletal complexity in the fungal kingdom
Abstract
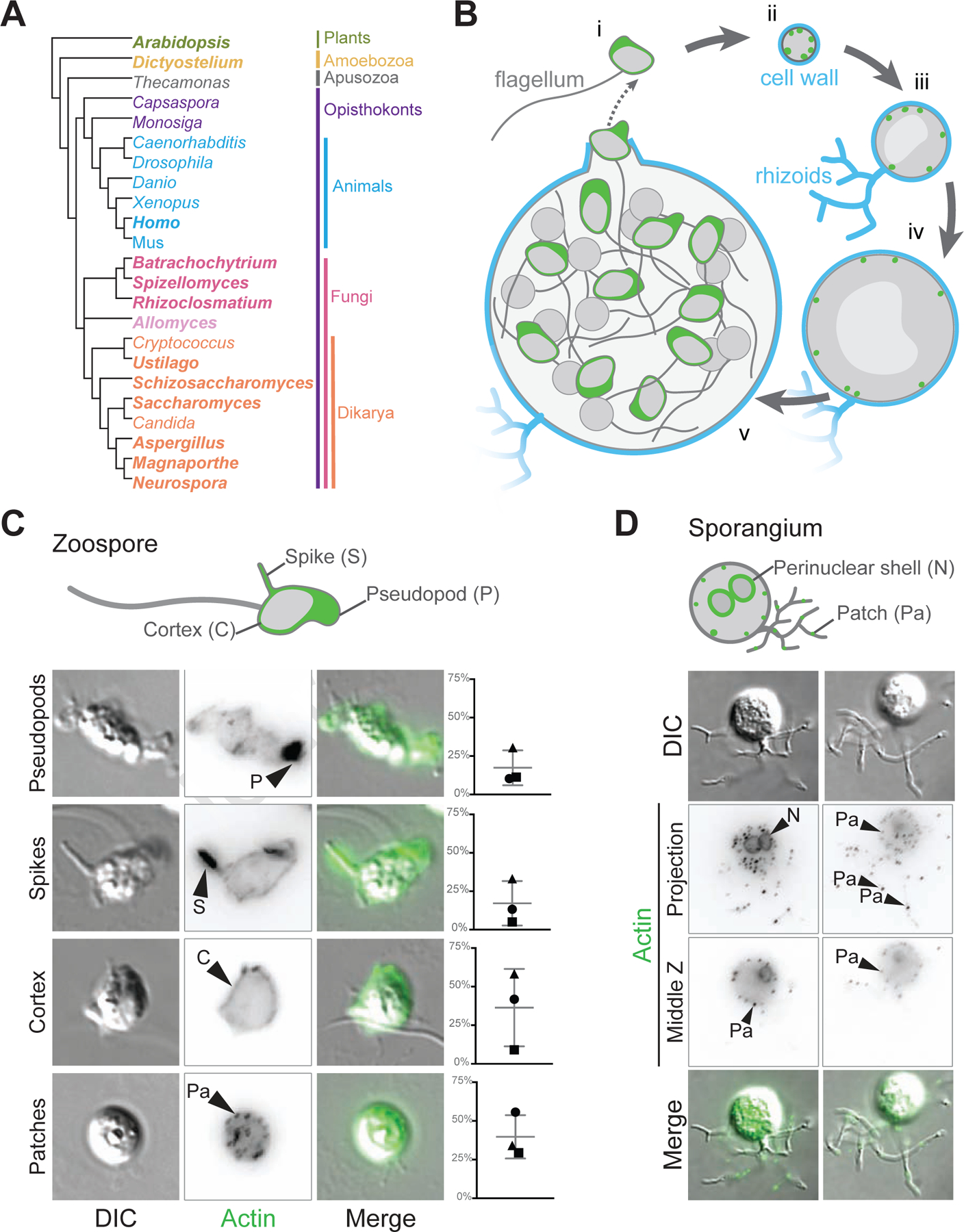
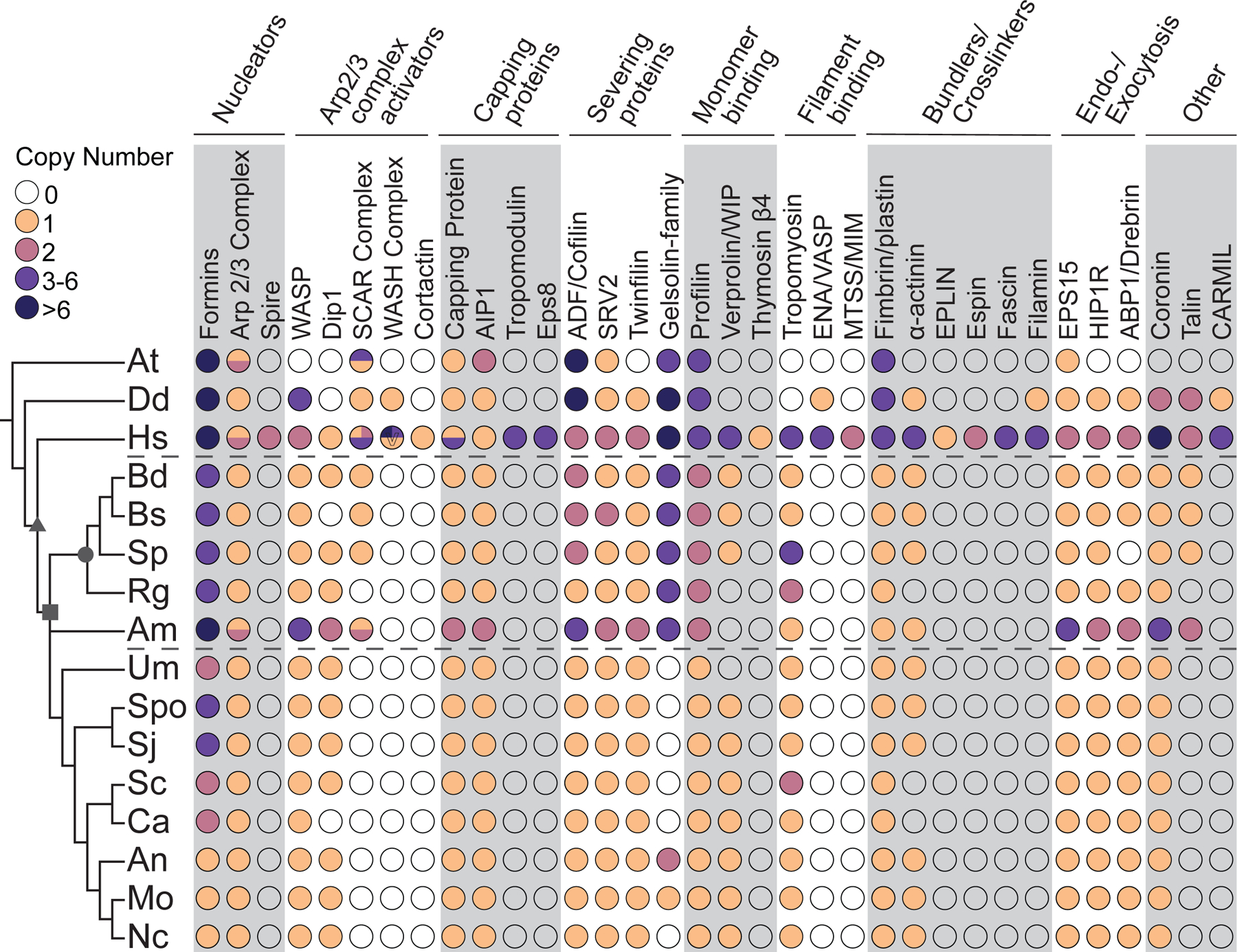
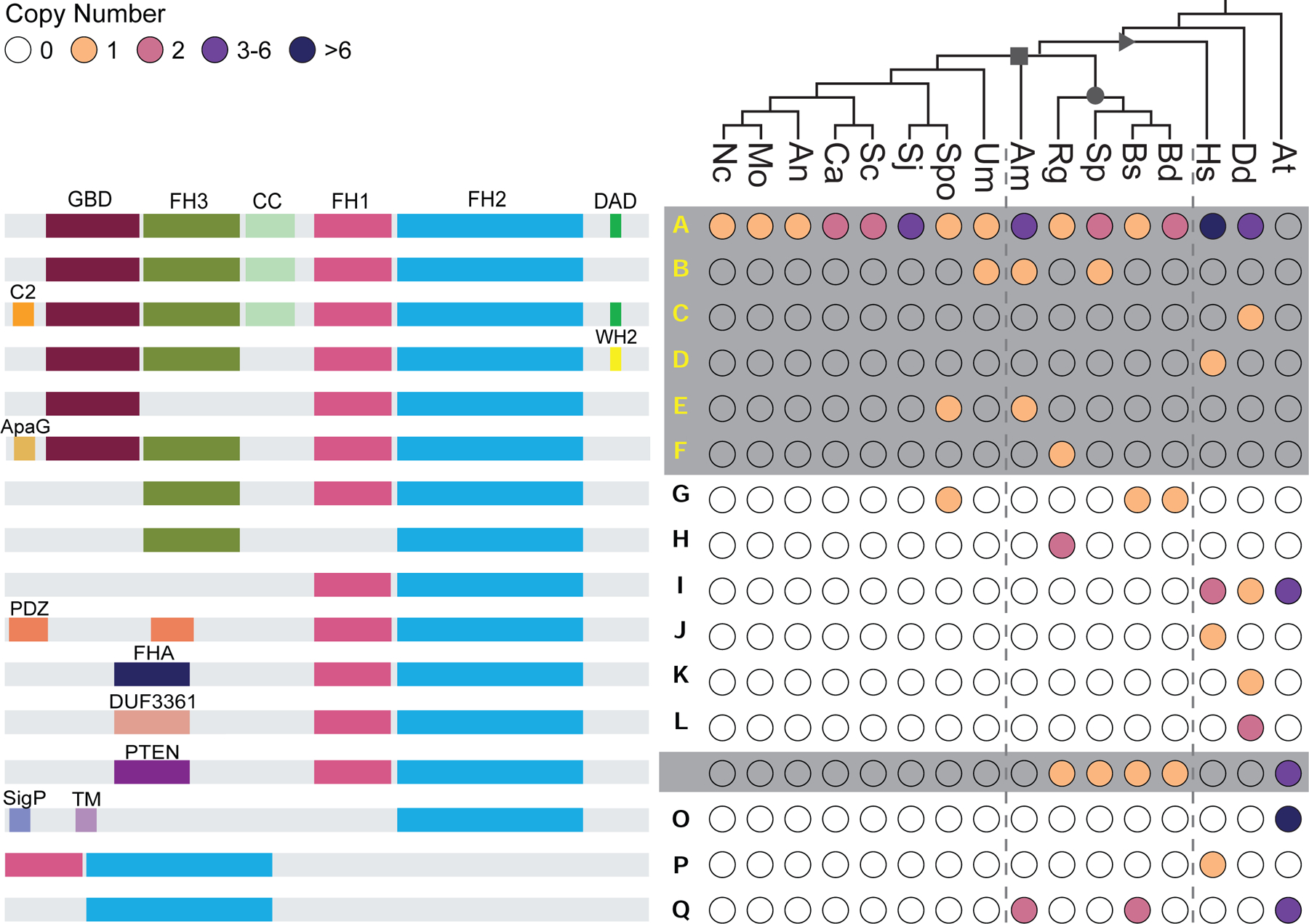
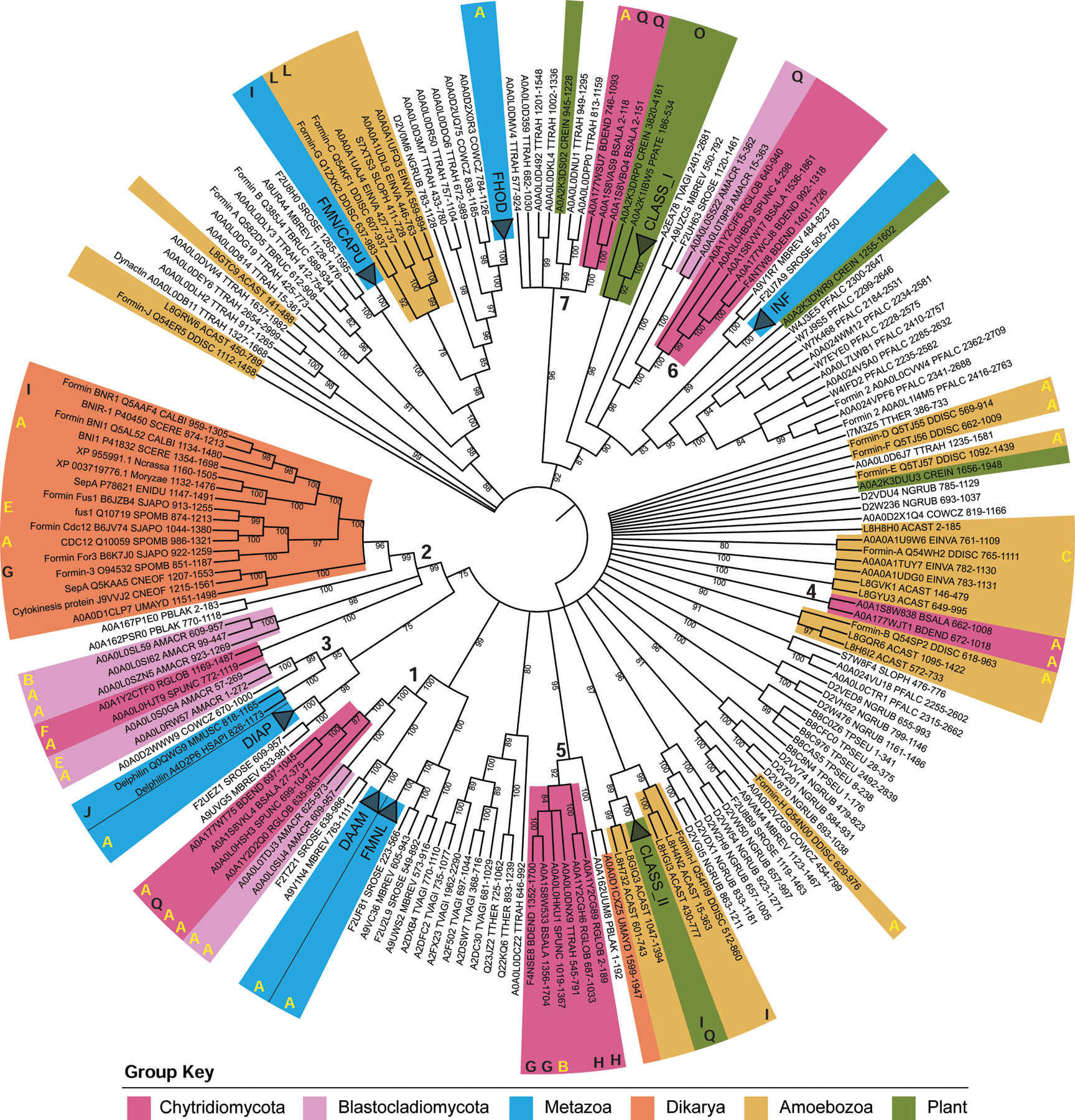
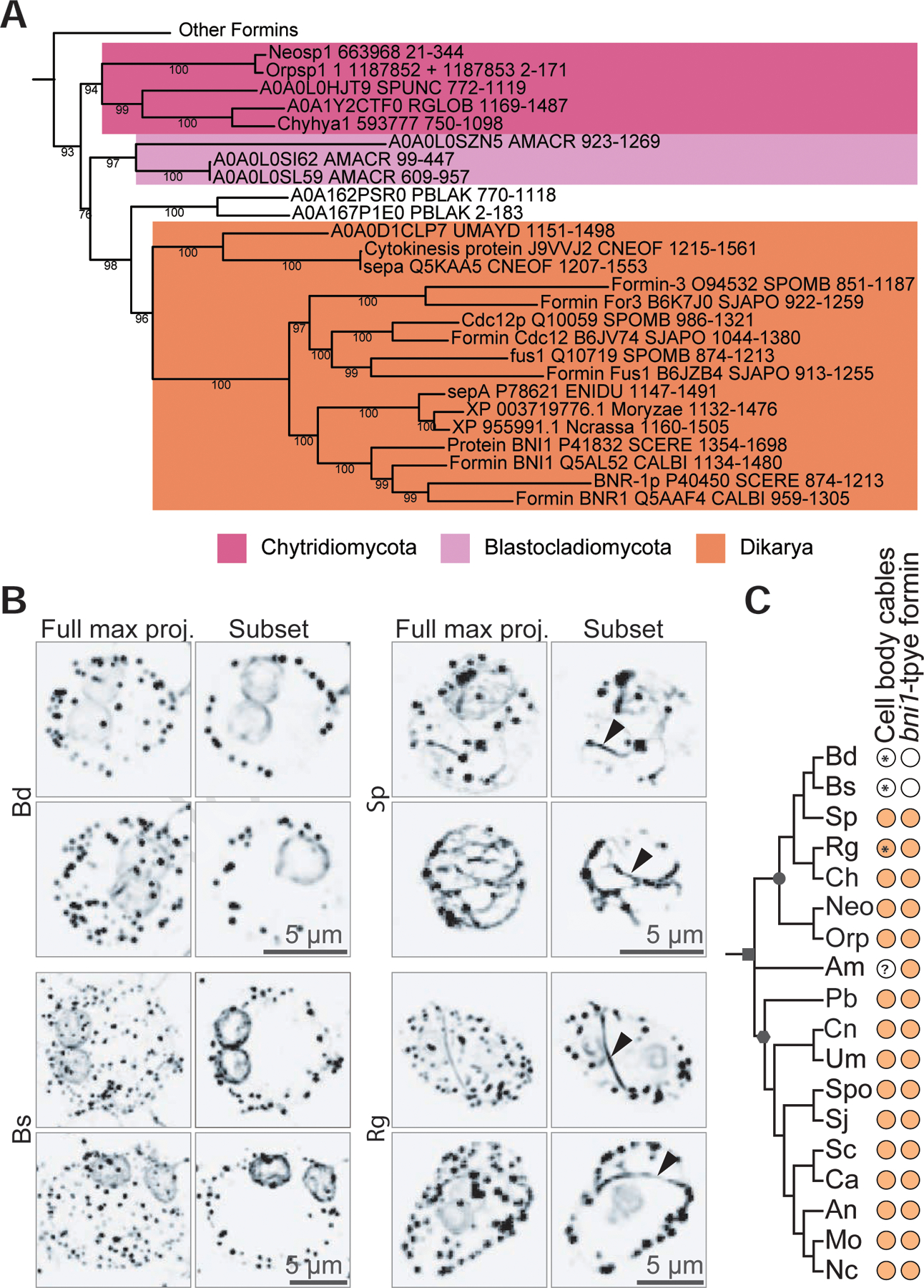
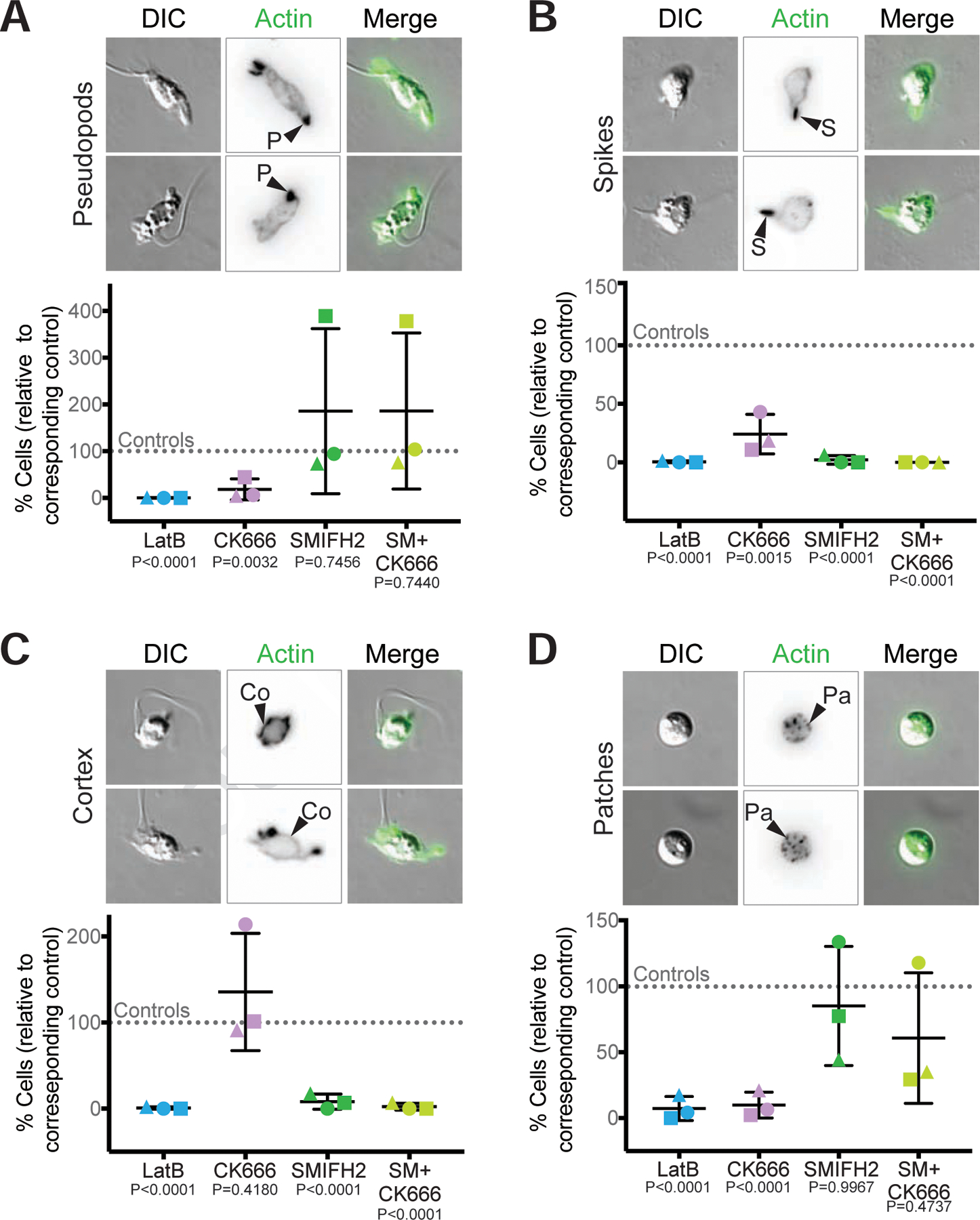
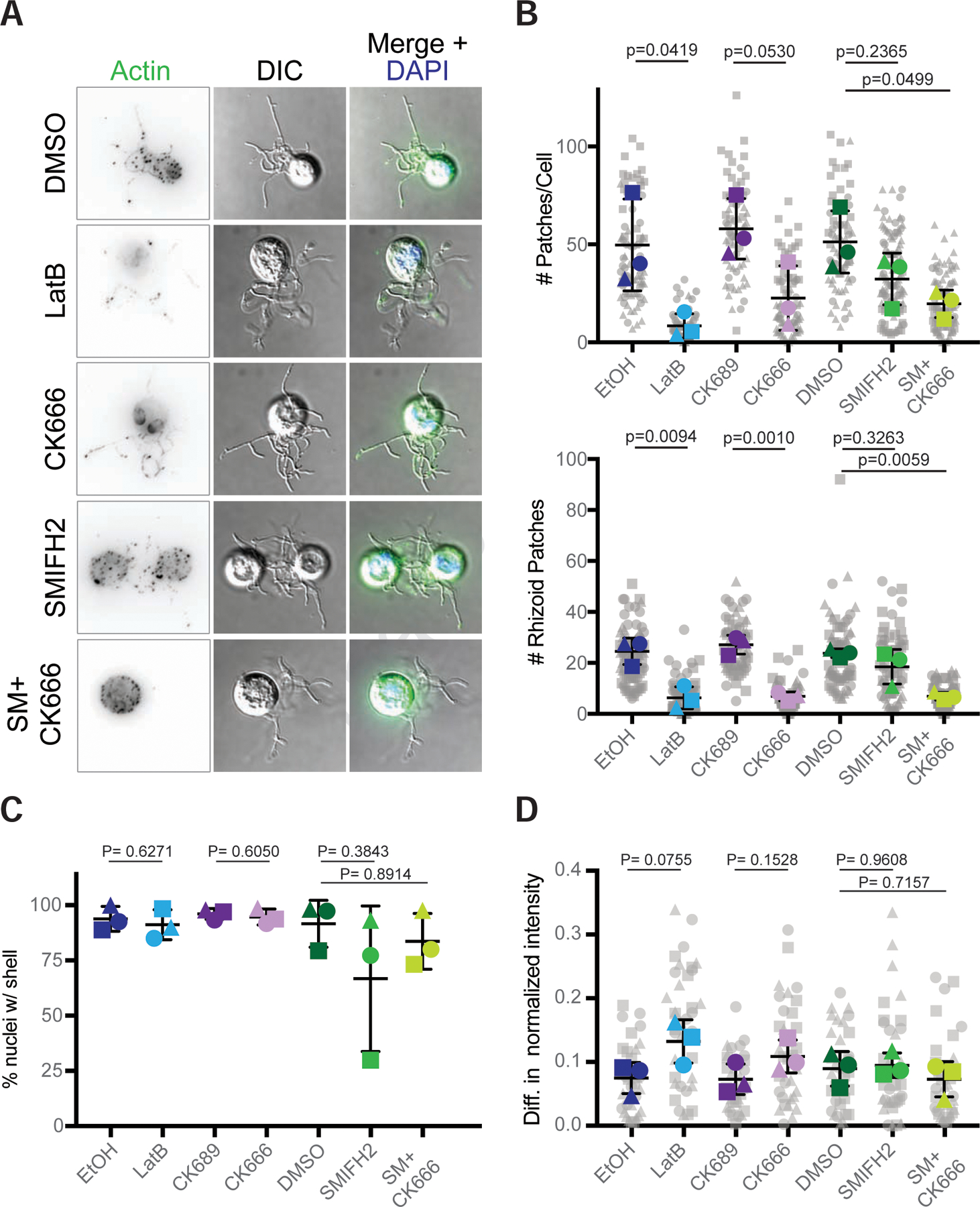
Cells from across the eukaryotic tree use actin polymer networks for a wide variety of functions, including endocytosis, cytokinesis, and cell migration. Despite this functional conservation, the actin cytoskeleton has undergone significant diversification, highlighted by the differences in the actin networks of mammalian cells and yeast. Chytrid fungi diverged before the emergence of the Dikarya (multicellular fungi and yeast) and therefore provide a unique opportunity to study actin cytoskeletal evolution. Chytrids have two life stages: zoospore cells that can swim with a flagellum and sessile sporangial cells that, like multicellular fungi, are encased in a chitinous cell wall. Here, we show that zoospores of the amphibian-killing chytrid Batrachochytrium dendrobatidis (Bd) build dynamic actin structures resembling those of animal cells, including an actin cortex, pseudopods, and filopodia-like spikes. In contrast, Bd sporangia assemble perinuclear actin shells and actin patches similar to those of yeast. The use of specific small-molecule inhibitors indicate that nearly all of Bd's actin structures are dynamic and use distinct nucleators: although pseudopods and actin patches are Arp2/3 dependent, the actin cortex appears formin dependent and actin spikes require both nucleators. Our analysis of multiple chytrid genomes reveals actin regulators and myosin motors found in animals, but not dikaryotic fungi, as well as fungal-specific components. The presence of animal- and yeast-like actin cytoskeletal components in the genome combined with the intermediate actin phenotypes in Bd suggests that the simplicity of the yeast cytoskeleton may be due to evolutionary loss.
Keywords: Batrachochytrium dendrobatidis; actin; chytrid; cytoskeleton; development; evolution; formin; fungi; motility; myosin.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Blanchoin L, Boujemaa-Paterski R, Sykes C, and Plastino J (2014). Actin dynamics, architecture, and mechanics in cell motility. Physiol. Rev 94, 235–263. - PubMed
-
- Velle KB, and Fritz-Laylin LK (2019). Diversity and evolution of actin-dependent phenotypes. Curr. Opin. Genet. Dev 58–59, 40–48. - PubMed
-
- Gallop JL (2020). Filopodia and their links with membrane traffic and cell adhesion. Semin. Cell Dev. Biol 102, 81–89. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
