

Interhemispheric transfer of working memories
- PMID: 33561399
- PMCID: PMC9134350
- DOI: 10.1016/j.neuron.2021.01.016
Interhemispheric transfer of working memories
Abstract
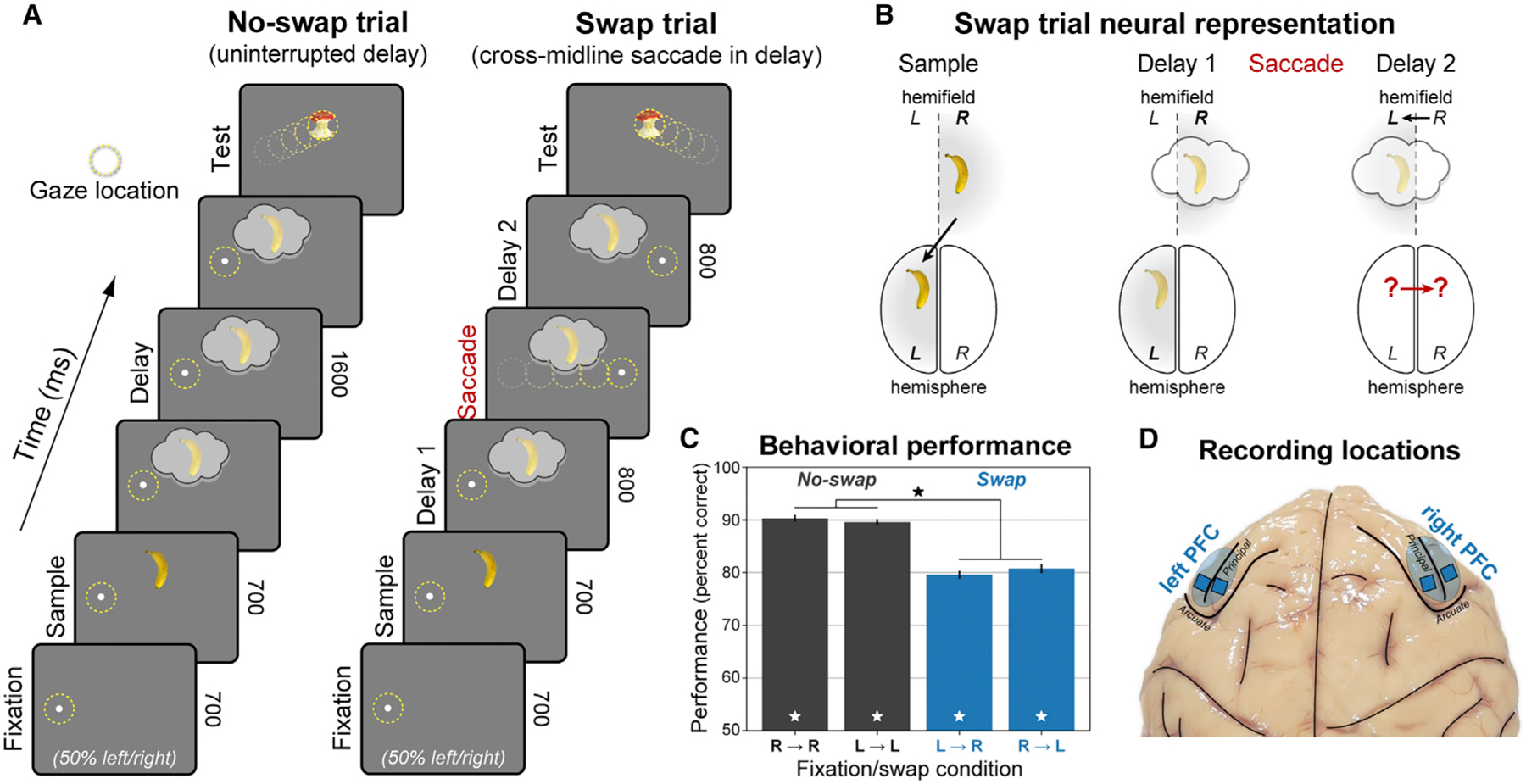
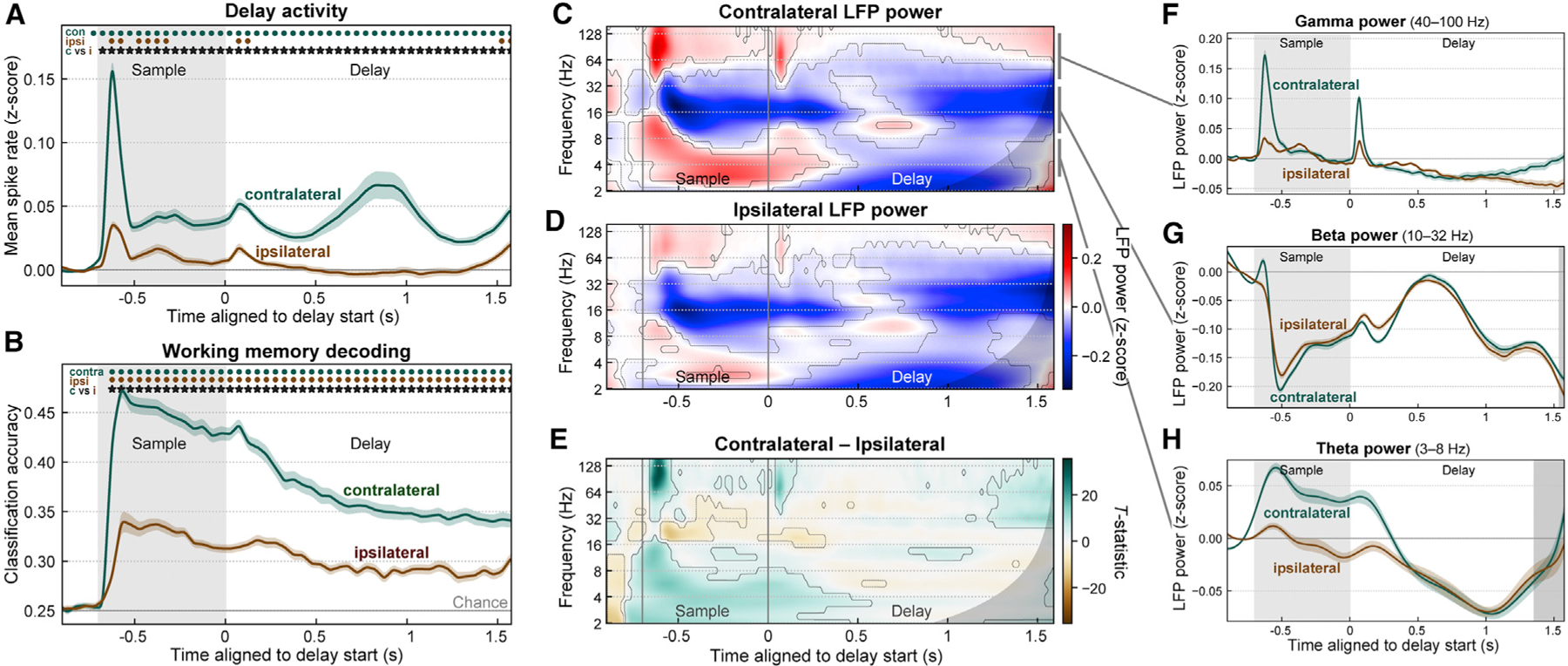
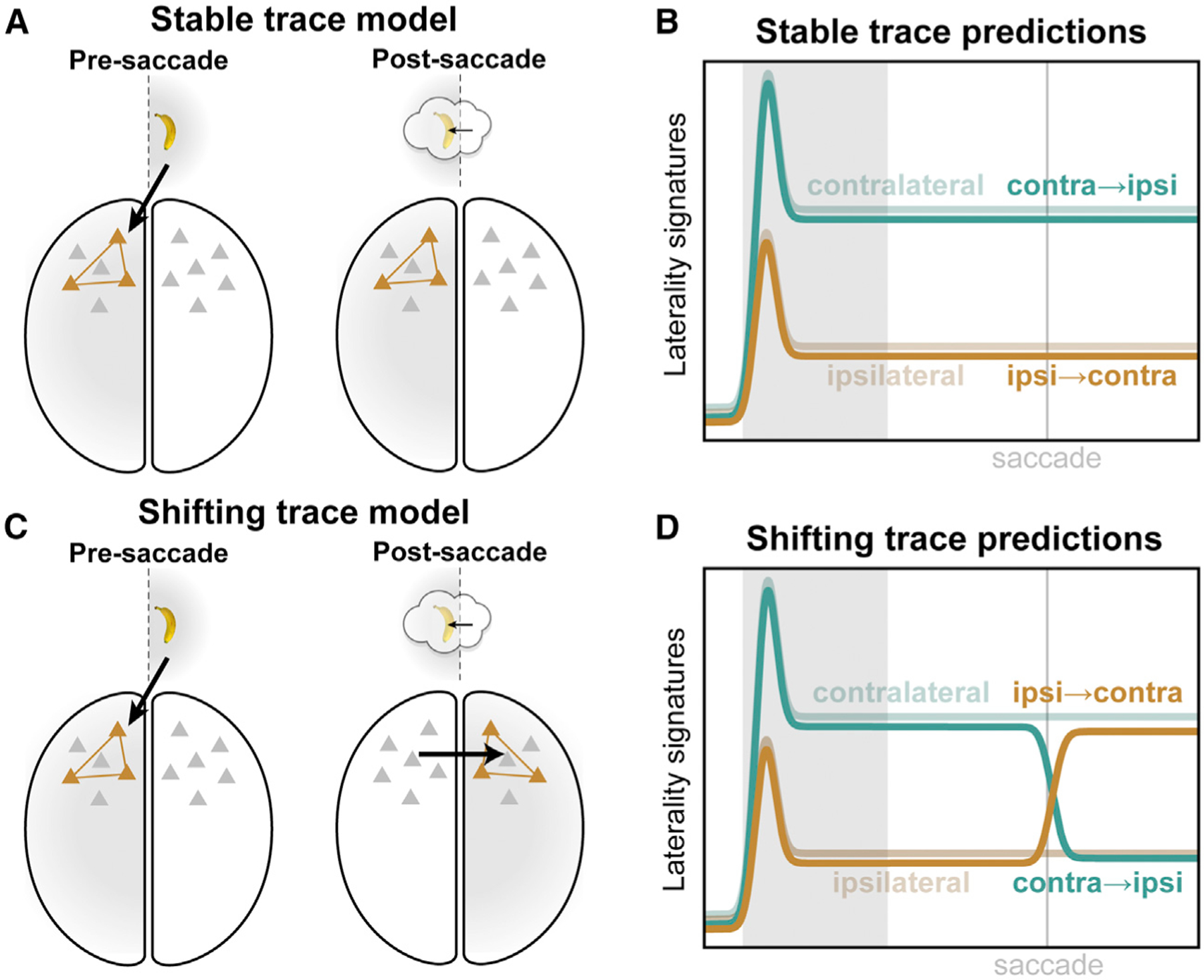
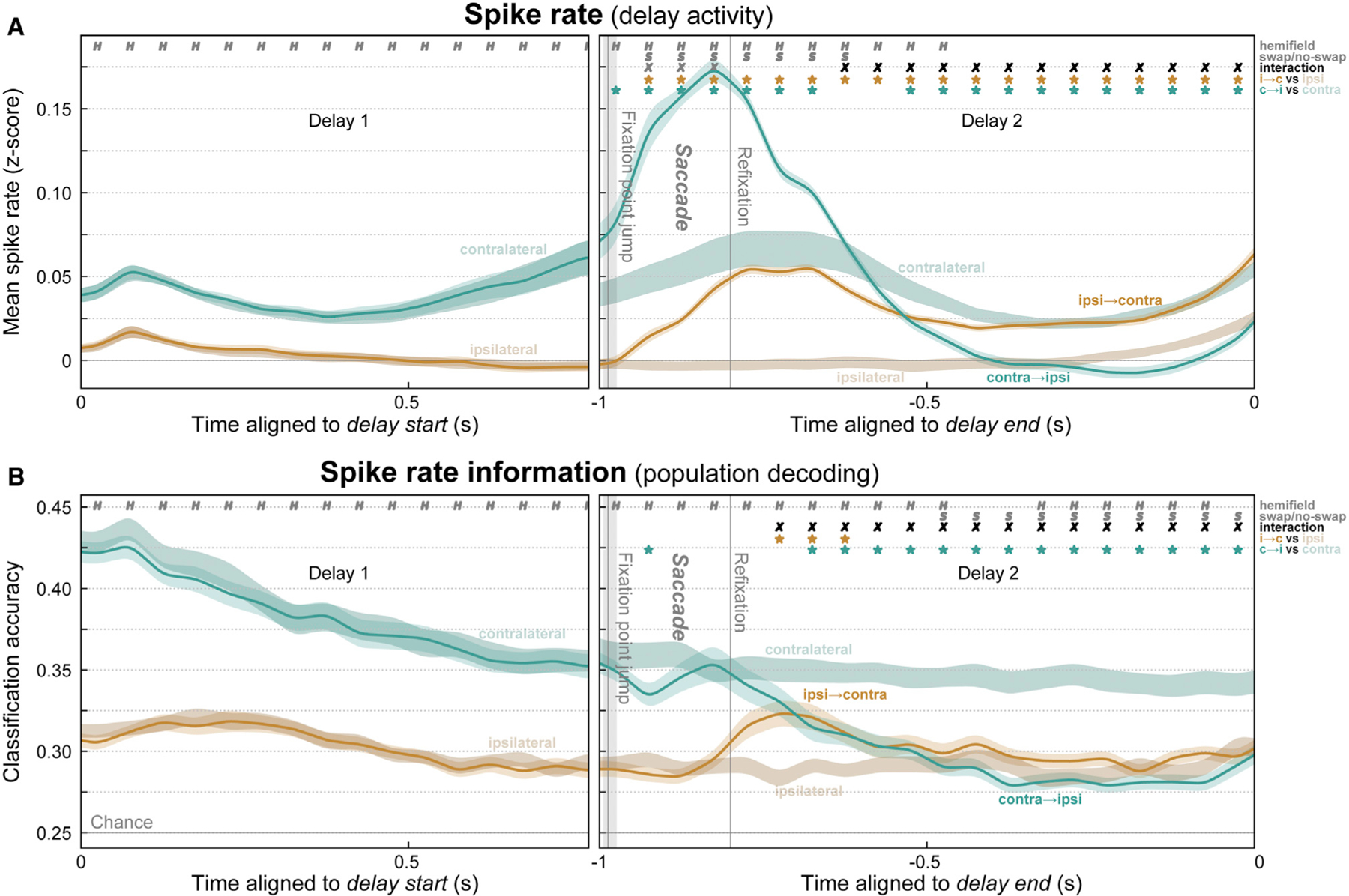
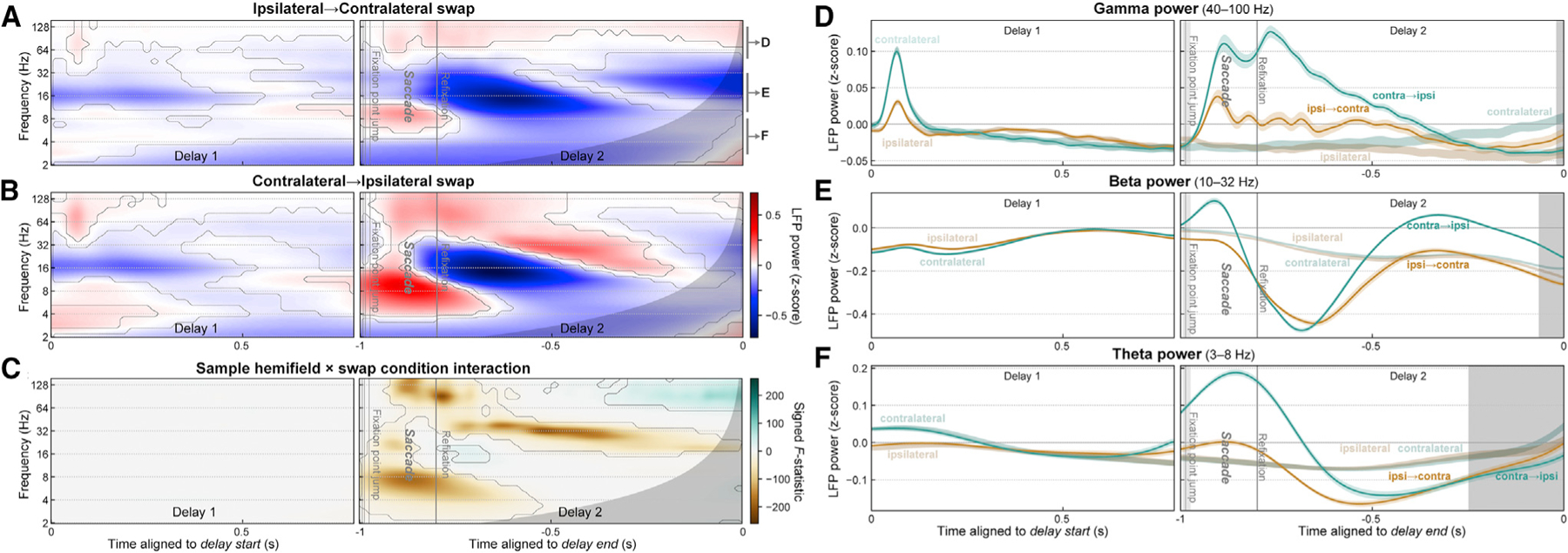
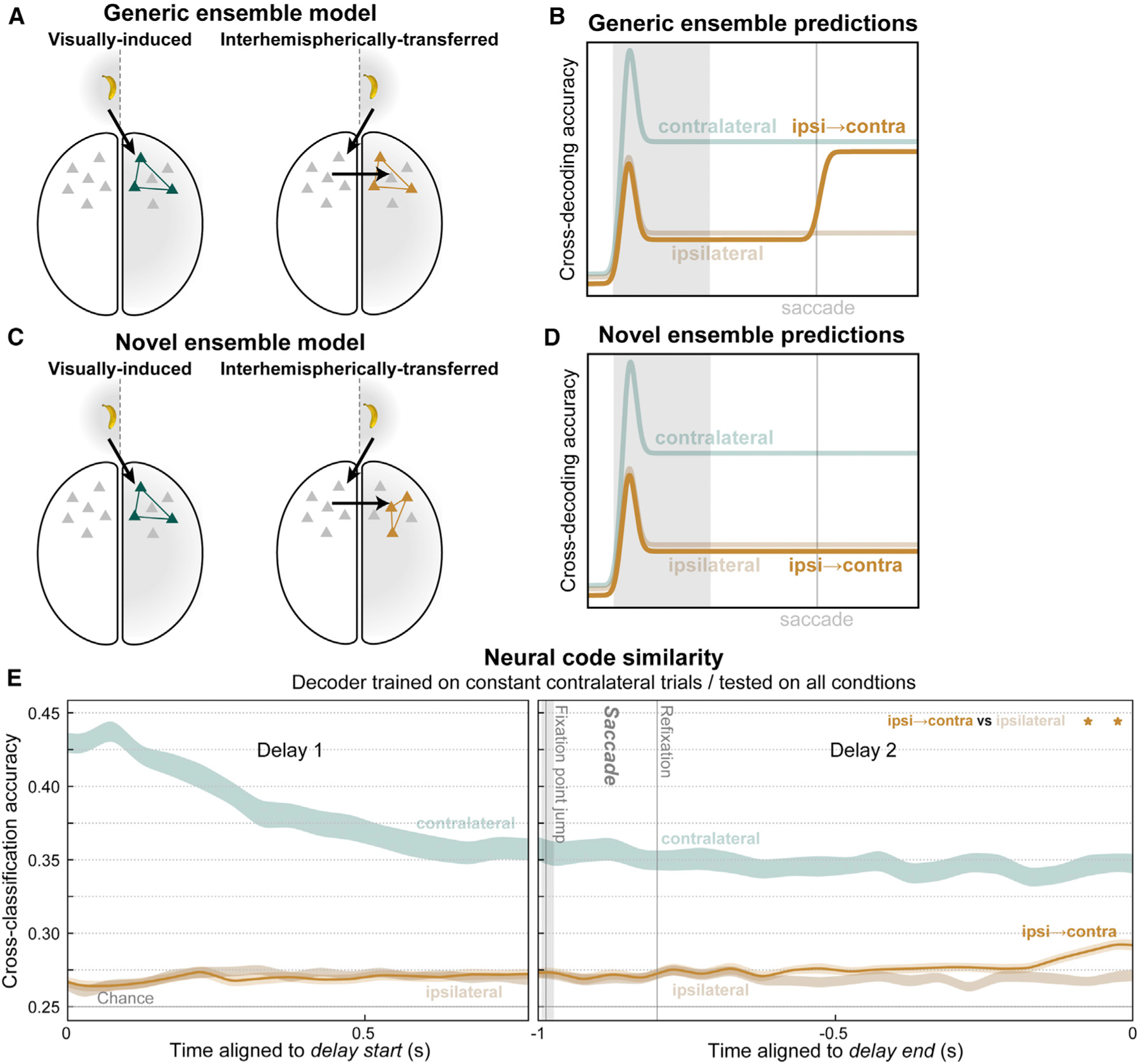
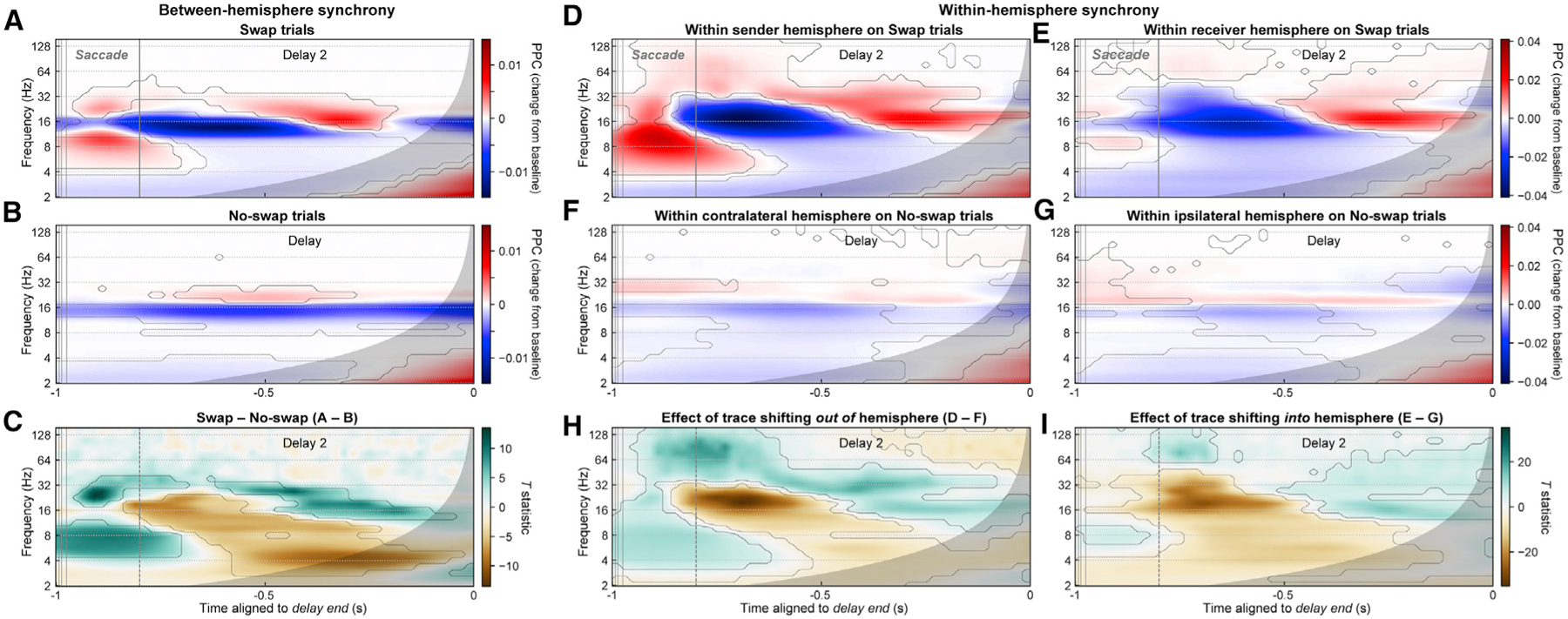
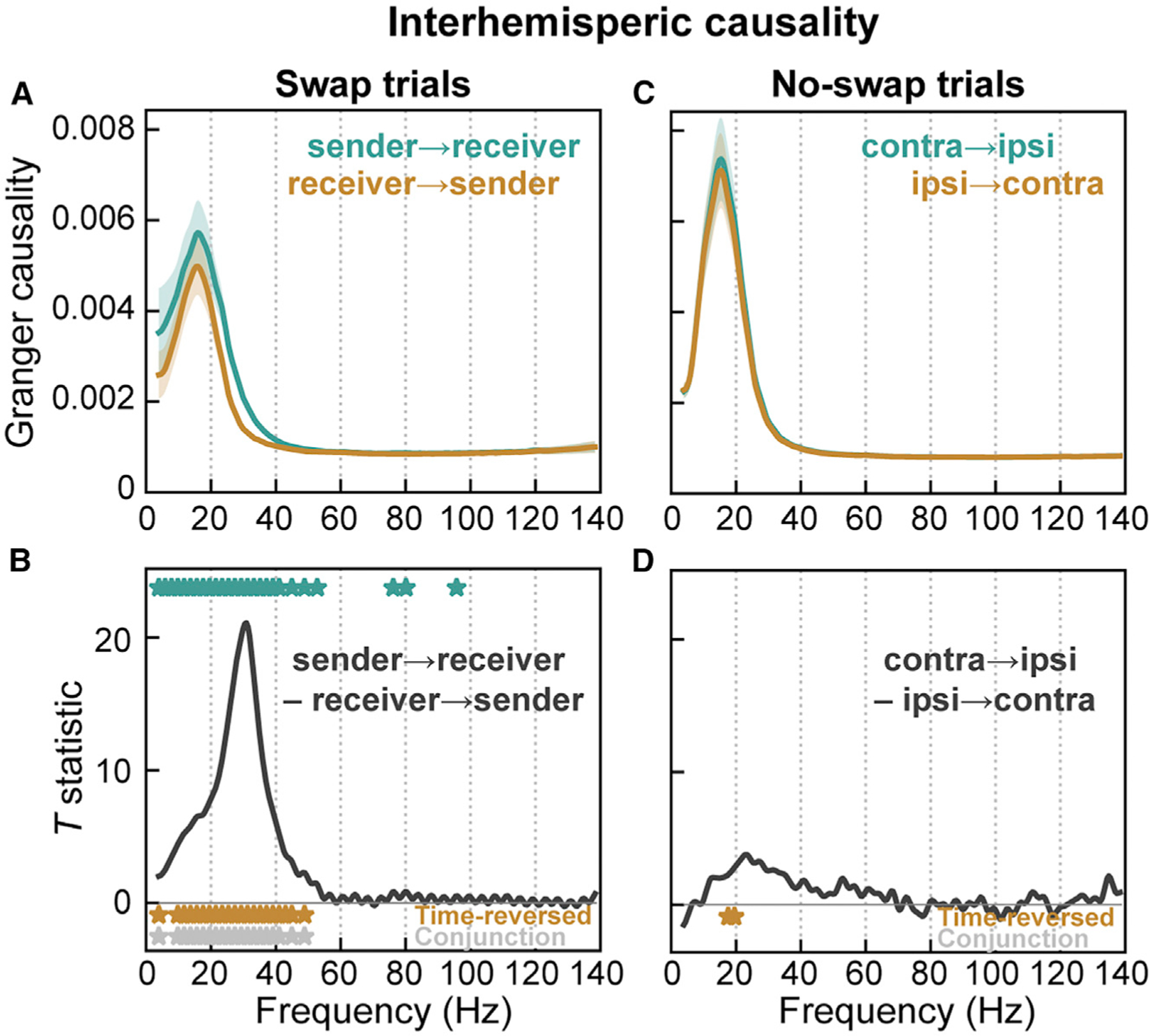
Visual working memory (WM) storage is largely independent between the left and right visual hemifields/cerebral hemispheres, yet somehow WM feels seamless. We studied how WM is integrated across hemifields by recording neural activity bilaterally from lateral prefrontal cortex. An instructed saccade during the WM delay shifted the remembered location from one hemifield to the other. Before the shift, spike rates and oscillatory power showed clear signatures of memory laterality. After the shift, the lateralization inverted, consistent with transfer of the memory trace from one hemisphere to the other. Transferred traces initially used different neural ensembles from feedforward-induced ones, but they converged at the end of the delay. Around the time of transfer, synchrony between the two prefrontal hemispheres peaked in theta and beta frequencies, with a directionality consistent with memory trace transfer. This illustrates how dynamics between the two cortical hemispheres can stitch together WM traces across visual hemifields.
Keywords: cognition; interhemispheric; neural synchrony; prefrontal cortex; working memory.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Hemispheres in harmony.Neuron. 2021 Mar 17;109(6):916-917. doi: 10.1016/j.neuron.2021.02.026. Neuron. 2021. PMID: 33735614
References
-
- Alvarez GA, and Cavanagh P (2005). Independent resources for attentional tracking in the left and right visual hemifields. Psychol. Sci 16, 637–643. - PubMed
-
- Barbas H, and Pandya DN (1984). Topography of commissural fibers of the prefrontal cortex in the rhesus monkey. Exp. Brain Res 55, 187–191. - PubMed
-
- Bastos AM, Vezoli J, Bosman CA, Schoffelen J-M, Oostenveld R, Dowdall JR, De Weerd P, Kennedy H, and Fries P (2015). Visual areas exert feedforward and feedback influences through distinct frequency channels. Neuron 85, 390–401. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
