

Target Score-A Proteomics Data Selection Tool Applied to Esophageal Cancer Identifies GLUT1-Sialyl Tn Glycoforms as Biomarkers of Cancer Aggressiveness
- PMID: 33562270
- PMCID: PMC7915893
- DOI: 10.3390/ijms22041664
Target Score-A Proteomics Data Selection Tool Applied to Esophageal Cancer Identifies GLUT1-Sialyl Tn Glycoforms as Biomarkers of Cancer Aggressiveness
Abstract
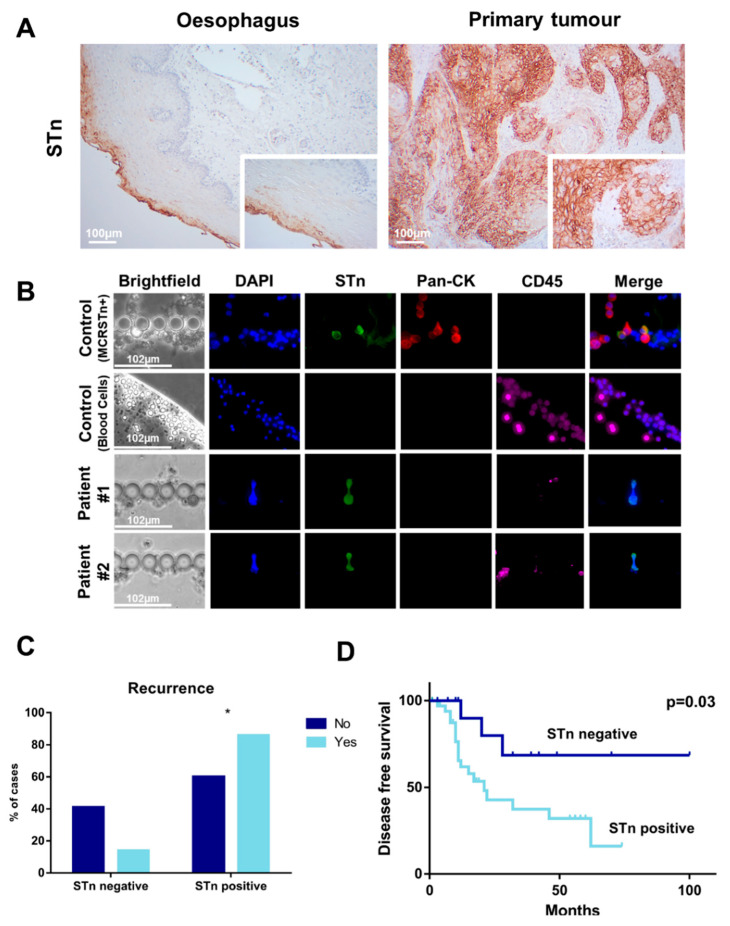
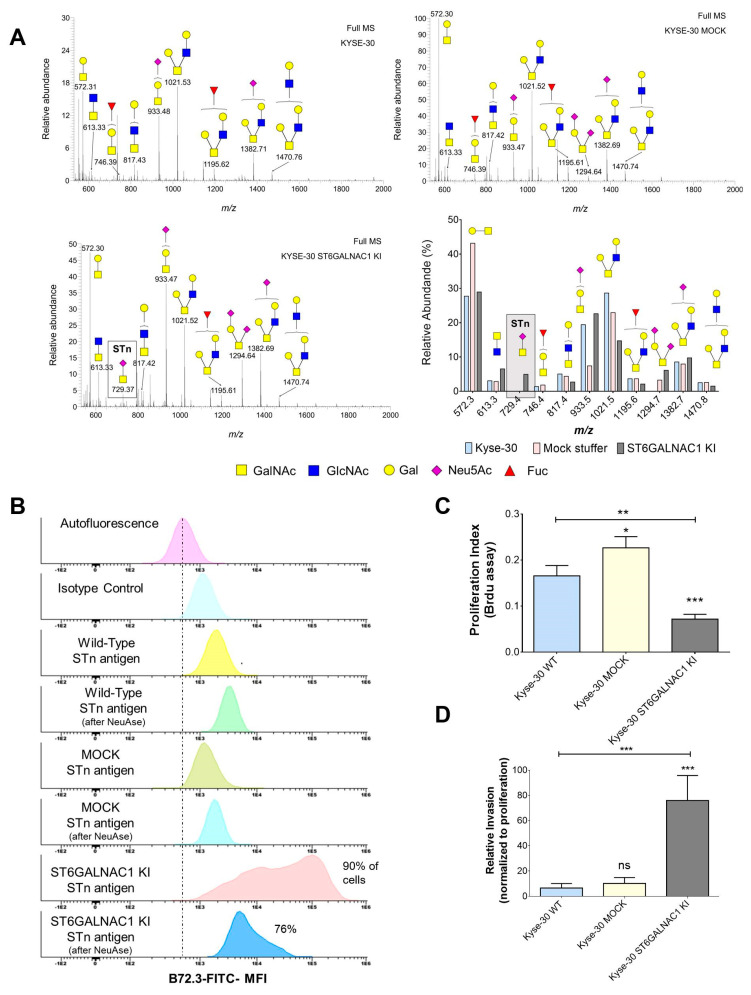
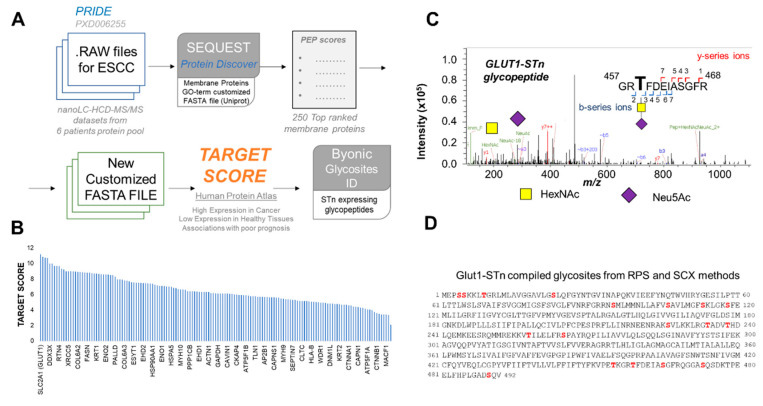
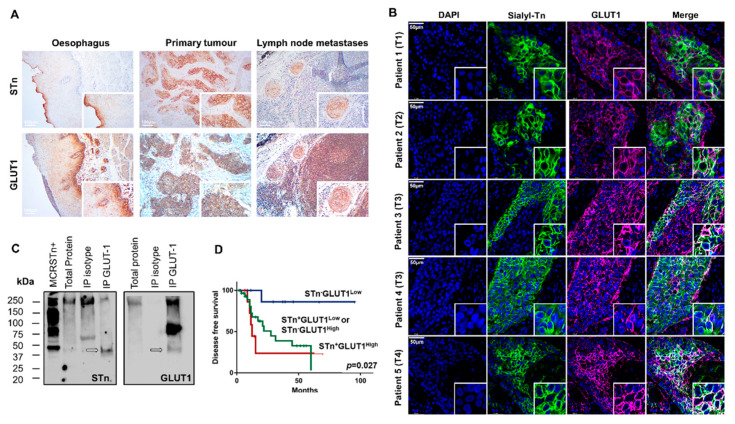
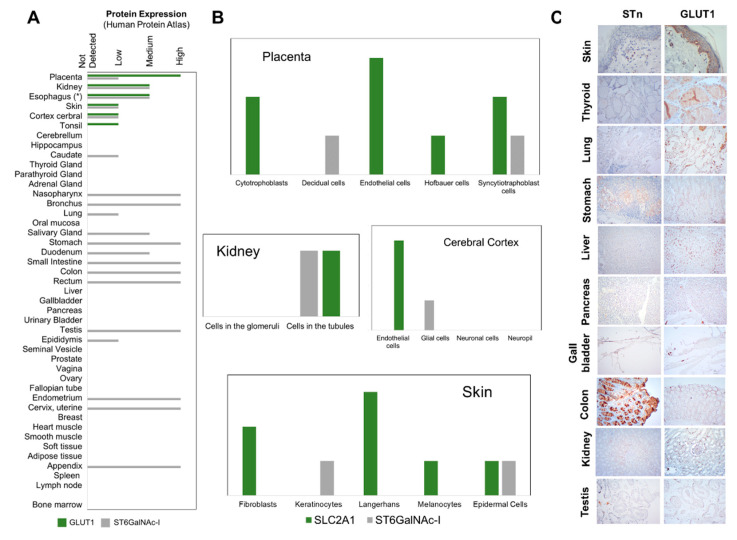
Esophageal cancer (EC) is a life-threatening disease, demanding the discovery of new biomarkers and molecular targets for precision oncology. Aberrantly glycosylated proteins hold tremendous potential towards this objective. In the current study, a series of esophageal squamous cell carcinomas (ESCC) and EC-derived circulating tumor cells (CTCs) were screened by immunoassays for the sialyl-Tn (STn) antigen, a glycan rarely expressed in healthy tissues and widely observed in aggressive gastrointestinal cancers. An ESCC cell model was glycoengineered to express STn and characterized in relation to cell proliferation and invasion in vitro. STn was found to be widely present in ESCC (70% of tumors) and in CTCs in 20% of patients, being associated with general recurrence and reduced survival. Furthermore, STn expression in ESCC cells increased invasion in vitro, while reducing cancer cells proliferation. In parallel, an ESCC mass spectrometry-based proteomics dataset, obtained from the PRIDE database, was comprehensively interrogated for abnormally glycosylated proteins. Data integration with the Target Score, an algorithm developed in-house, pinpointed the glucose transporter type 1 (GLUT1) as a biomarker of poor prognosis. GLUT1-STn glycoproteoforms were latter identified in tumor tissues in patients facing worst prognosis. Furthermore, healthy human tissues analysis suggested that STn glycosylation provided cancer specificity to GLUT1. In conclusion, STn is a biomarker of worst prognosis in EC and GLUT1-STn glycoforms may be used to increase its specificity on the stratification and targeting of aggressive ESCC forms.
Keywords: bioinformatics; cancer biomarkers; circulating tumors cells; esophageal cancer; glycomics; glycoproteomics.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
MeSH terms
Substances
Grants and funding
- SFRH/BD/111242/2015/Fundação para a Ciência e a Tecnologia
- SFRH/BD/146500/2019/Fundação para a Ciência e a Tecnologia
- SFRH/BD/142479/2018/Fundação para a Ciência e a Tecnologia
- SFRH/BD/105355/2014/Fundação para a Ciência e a Tecnologia
- CEECIND/03186/2017/Fundação para a Ciência e a Tecnologia
- PEst-OE/SAU/UI0776/201/Fundação para a Ciência e a Tecnologia
- CI-IPOP-29-2020/Instituto Português de Oncologia do Porto
- CI-IPOP-58-2020/Instituto Português de Oncologia do Porto
- CI-IPOP-Proj.70-bolsa2019-GPTE/Instituto Português de Oncologia do Porto
- UIDB/50006/2020/Fundação para a Ciência e a Tecnologia
- DSAIPA/DS/0042/2018/Fundação para a Ciência e a Tecnologia
- NORTE-01-0145-FEDER-000027/European Regional Development Fund
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
