

CD20 as a gatekeeper of the resting state of human B cells
- PMID: 33563755
- PMCID: PMC7896350
- DOI: 10.1073/pnas.2021342118
CD20 as a gatekeeper of the resting state of human B cells
Abstract
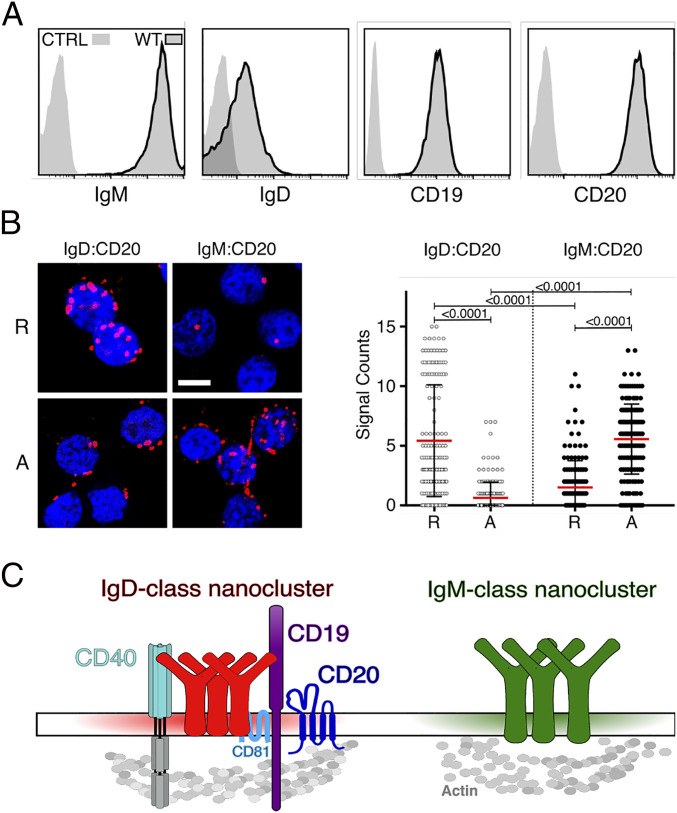
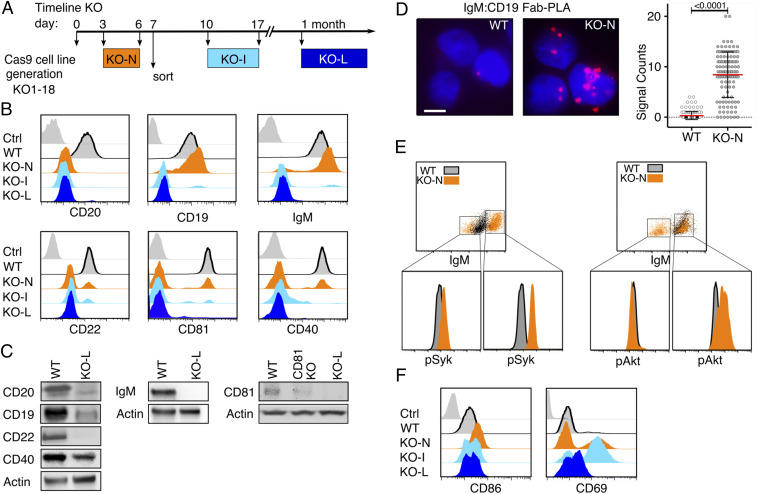
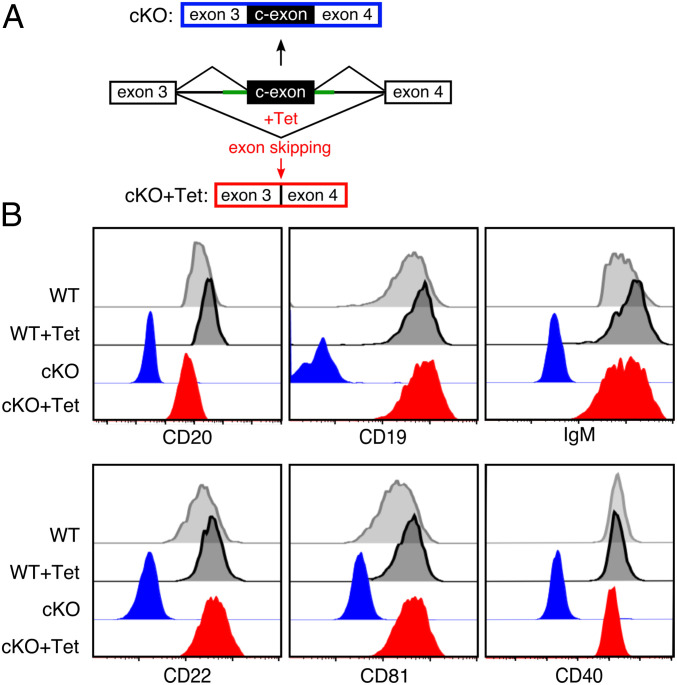
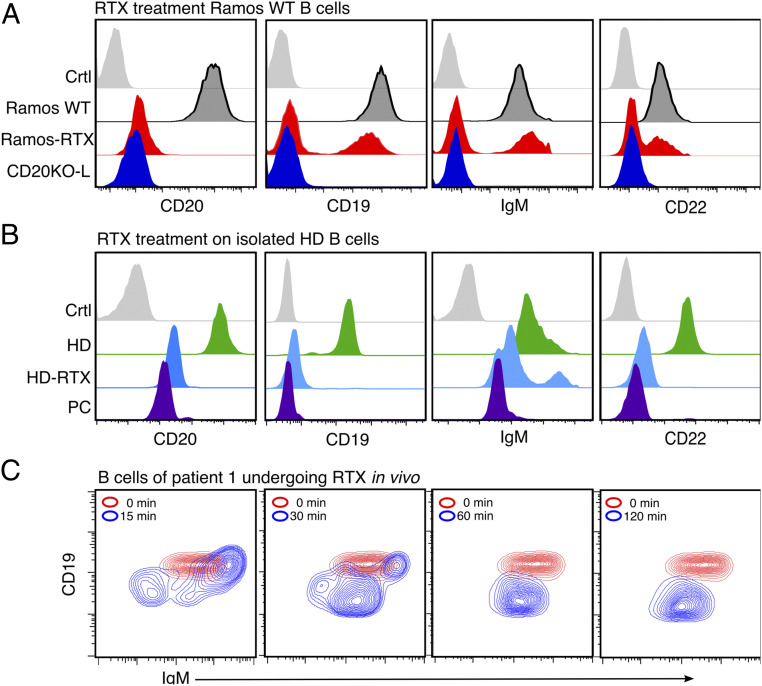
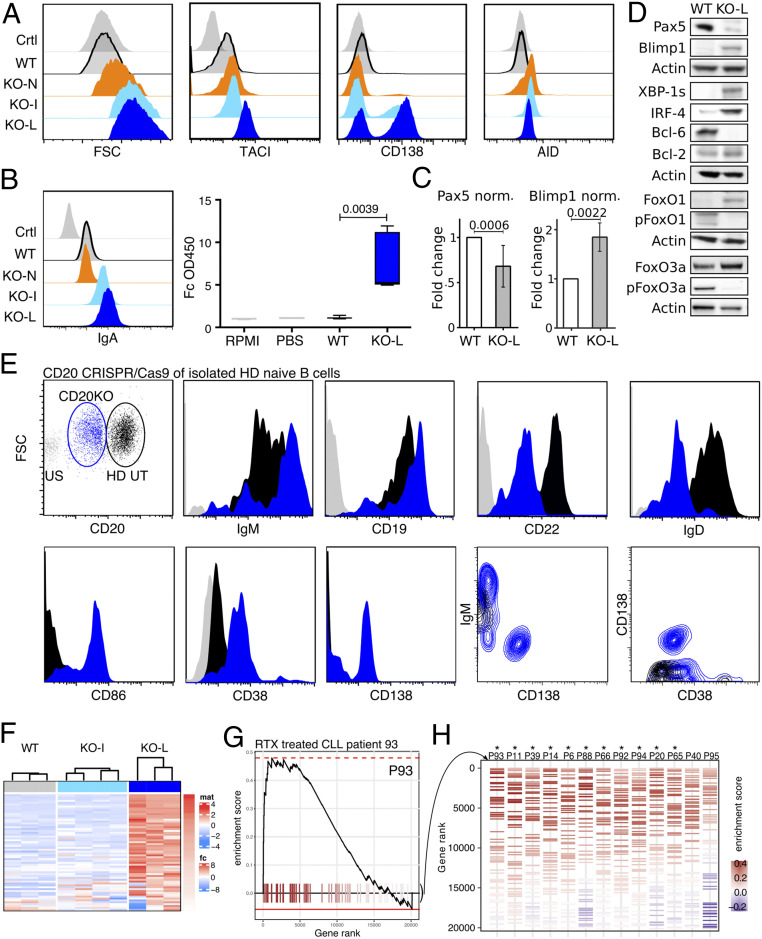
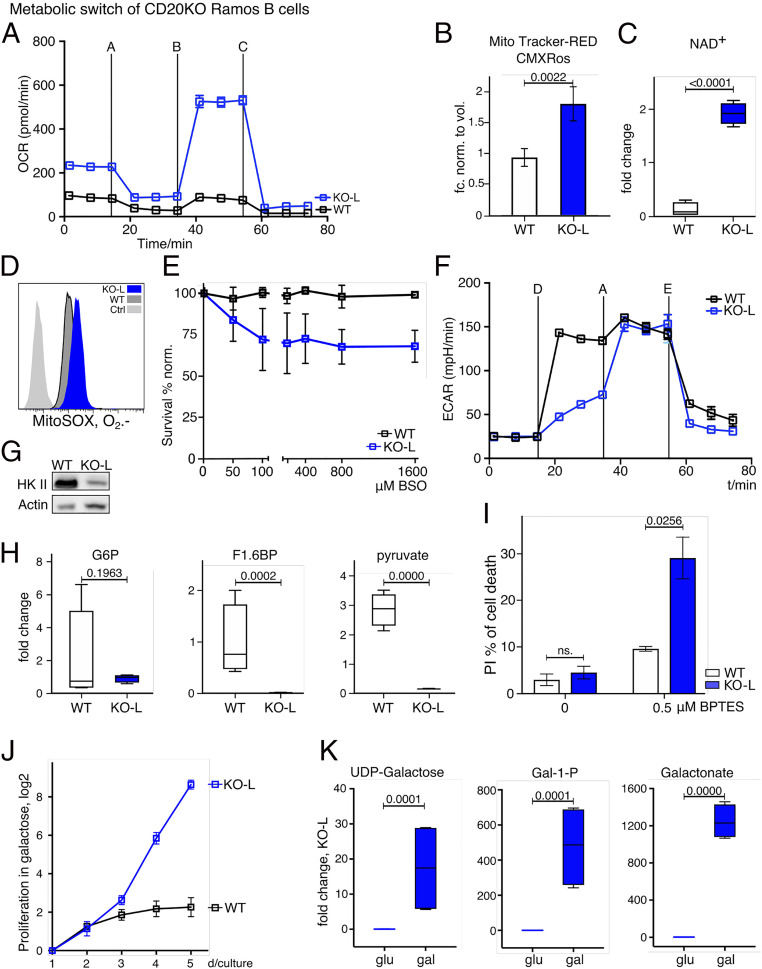
CD20 is a B cell-specific membrane protein and represents an attractive target for therapeutic antibodies. Despite widespread usage of anti-CD20 antibodies for B cell depletion therapies, the biological function of their target remains unclear. Here, we demonstrate that CD20 controls the nanoscale organization of receptors on the surface of resting B lymphocytes. CRISPR/Cas9-mediated ablation of CD20 in resting B cells resulted in relocalization and interaction of the IgM-class B cell antigen receptor with the coreceptor CD19. This receptor rearrangement led to a transient activation of B cells, accompanied by the internalization of many B cell surface marker proteins. Reexpression of CD20 restored the expression of the B cell surface proteins and the resting state of Ramos B cells. Similarly, treatment of Ramos or naive human B cells with the anti-CD20 antibody rituximab induced nanoscale receptor rearrangements and transient B cell activation in vitro and in vivo. A departure from the resting B cell state followed by the loss of B cell identity of CD20-deficient Ramos B cells was accompanied by a PAX5 to BLIMP-1 transcriptional switch, metabolic reprogramming toward oxidative phosphorylation, and a shift toward plasma cell development. Thus, anti-CD20 engagement or the loss of CD20 disrupts membrane organization, profoundly altering the fate of human B cells.
Keywords: B lymphocyte; CD20; plasma cell; therapeutic antibody.
Copyright © 2021 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
CD19 and CD20 targeted vectors induce minimal activation of resting B lymphocytes.PLoS One. 2013 Nov 11;8(11):e79047. doi: 10.1371/journal.pone.0079047. eCollection 2013. PLoS One. 2013. PMID: 24244415 Free PMC article.
-
Enhancement of Rituximab-induced cell death by the physical association of CD20 with CD40 molecules on the cell surface.Int Immunol. 2014 Aug;26(8):451-65. doi: 10.1093/intimm/dxu046. Epub 2014 May 15. Int Immunol. 2014. PMID: 24894009
-
Defective calcium signaling and disrupted CD20-B-cell receptor dissociation in patients with common variable immunodeficiency disorders.J Allergy Clin Immunol. 2012 Mar;129(3):755-761.e7. doi: 10.1016/j.jaci.2011.10.020. Epub 2011 Nov 30. J Allergy Clin Immunol. 2012. PMID: 22130422
-
Functional role of lipid rafts in CD20 activity?Biochem Soc Symp. 2005;(72):165-75. doi: 10.1042/bss0720165. Biochem Soc Symp. 2005. PMID: 15649140 Review.
-
The Tumor Microenvironment Regulates CD19 and CD20 Immunotherapy for Lymphoma.Cancer J. 2015 Jul-Aug;21(4):351-6. doi: 10.1097/PPO.0000000000000137. Cancer J. 2015. PMID: 26222089 Review.
Cited by
-
Bayesian-based analysis of the causality between 731 immune cells and erectile dysfunction: a two-sample, bidirectional, and multivariable Mendelian randomization study.Sex Med. 2024 Sep 21;12(4):qfae062. doi: 10.1093/sexmed/qfae062. eCollection 2024 Aug. Sex Med. 2024. PMID: 39315306 Free PMC article.
-
Comparison of the blood immune repertoire with clinical features in chronic lymphocytic leukemia patients treated with chemoimmunotherapy or ibrutinib.Front Oncol. 2023 Dec 4;13:1302038. doi: 10.3389/fonc.2023.1302038. eCollection 2023. Front Oncol. 2023. PMID: 38111528 Free PMC article.
-
Learning cell identity in immunology, neuroscience, and cancer.Semin Immunopathol. 2023 Jan;45(1):3-16. doi: 10.1007/s00281-022-00976-y. Epub 2022 Dec 19. Semin Immunopathol. 2023. PMID: 36534139 Free PMC article. Review.
-
M2 Macrophage and Extracellular Matrix Genes Are Enriched in High-Activity Lichen Planopilaris.Dermatol Res Pract. 2025 May 29;2025:5545886. doi: 10.1155/drp/5545886. eCollection 2025. Dermatol Res Pract. 2025. PMID: 40475059 Free PMC article.
-
Targeting B cells and plasma cells in autoimmune diseases: From established treatments to novel therapeutic approaches.Eur J Immunol. 2023 Jan;53(1):e2149675. doi: 10.1002/eji.202149675. Epub 2022 Nov 16. Eur J Immunol. 2023. PMID: 36314264 Free PMC article. Review.
References
-
- Kosmas C., Stamatopoulos K., Stavroyianni N., Tsavaris N., Papadaki T., Anti-CD20-based therapy of B cell lymphoma: State of the art. Leukemia 16, 2004–2015 (2002). - PubMed
-
- Awasthi A., Rolland D. C. M., Ayello J.et al. , A comparative global phosphoproteomics analysis of obinutuzumab (GA101) versus rituximab (RTX) against RTX sensitive and resistant Burkitt lymphoma (BL) demonstrates differential phosphorylation of signaling pathway proteins after treatment. Oncotarget 8, 113895–113909 (2017). - PMC - PubMed
-
- Mok C. C., Current role of rituximab in systemic lupus erythematosus. Int. J. Rheum. Dis. 18, 154–163 (2015). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
