

The branched-chain amino acid aminotransferase TaBCAT1 modulates amino acid metabolism and positively regulates wheat rust susceptibility
- PMID: 33565586
- PMCID: PMC8254495
- DOI: 10.1093/plcell/koab049
The branched-chain amino acid aminotransferase TaBCAT1 modulates amino acid metabolism and positively regulates wheat rust susceptibility
Abstract
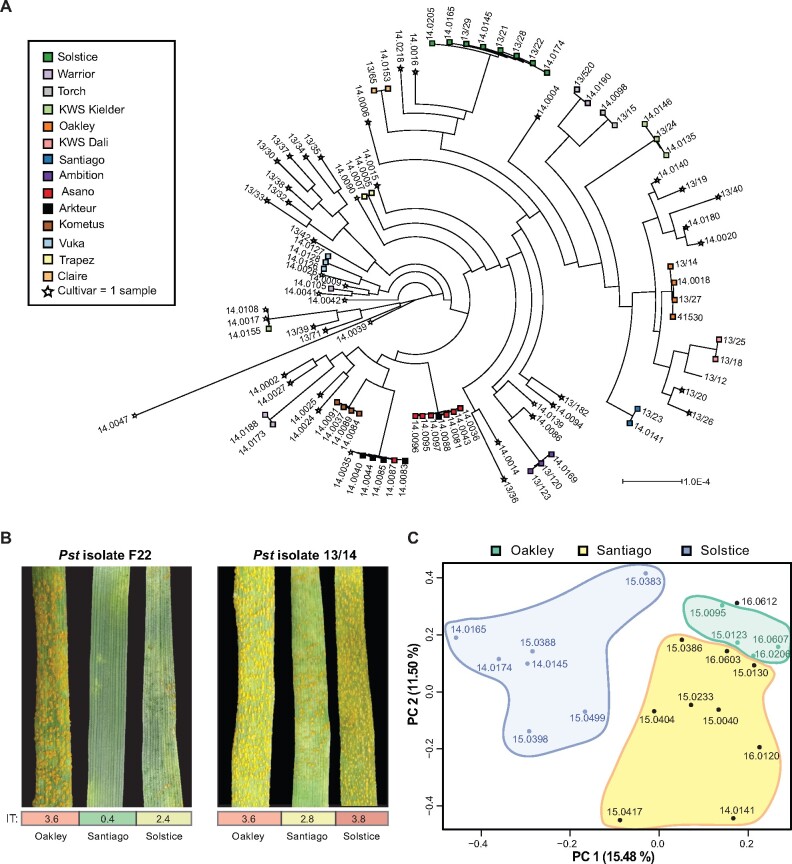
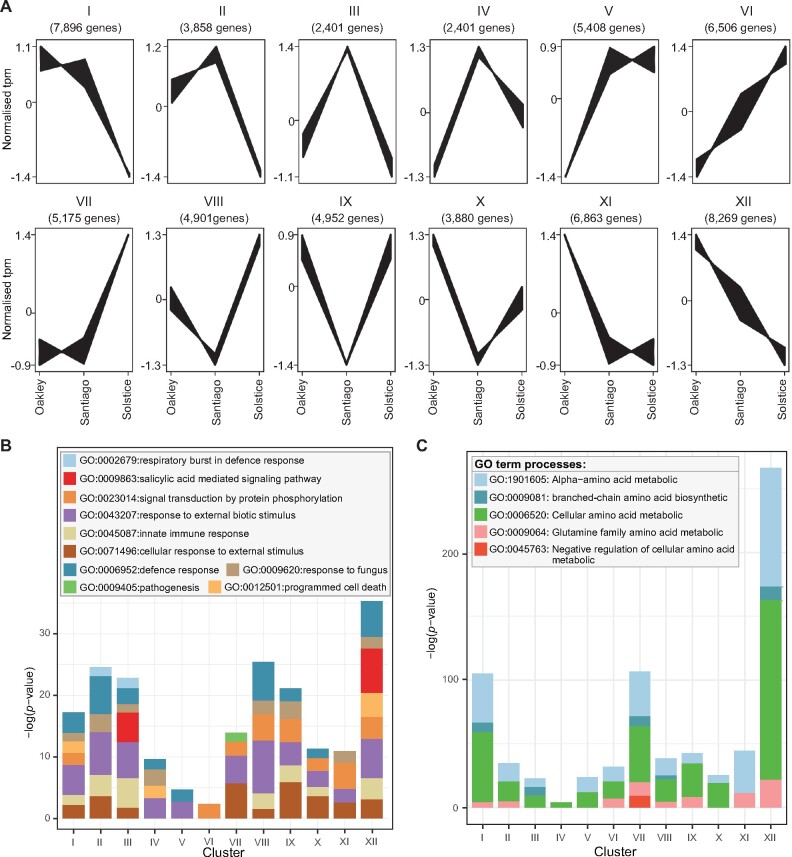
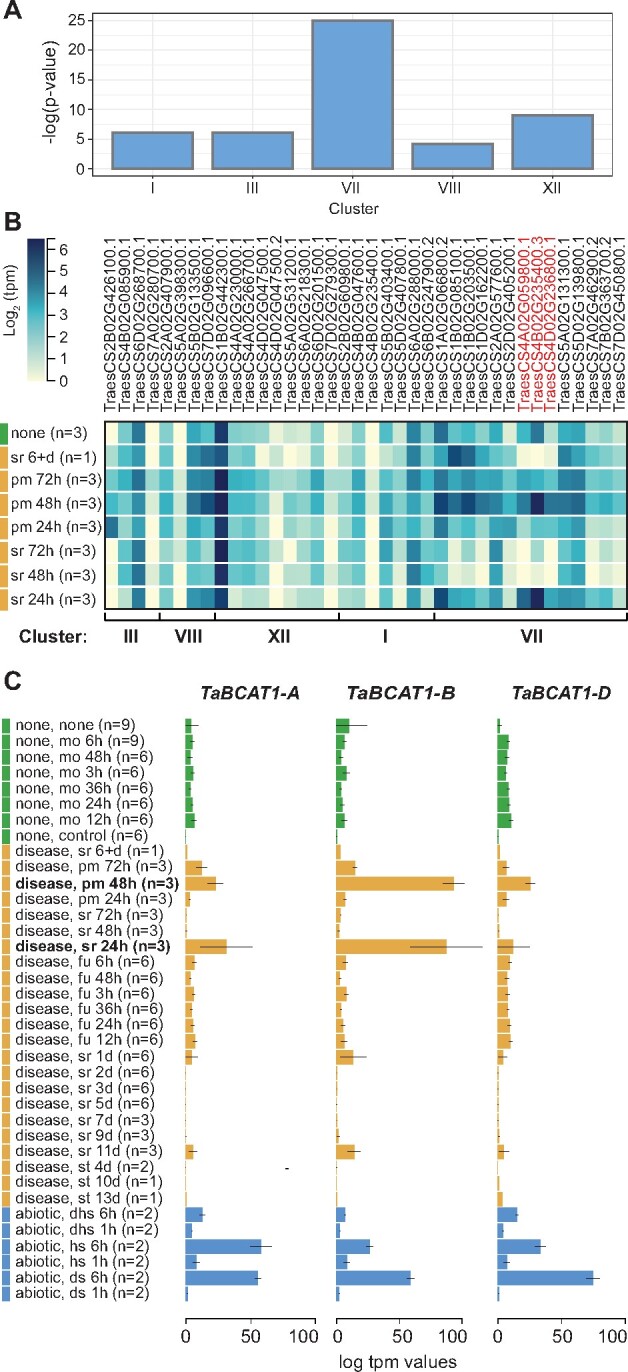
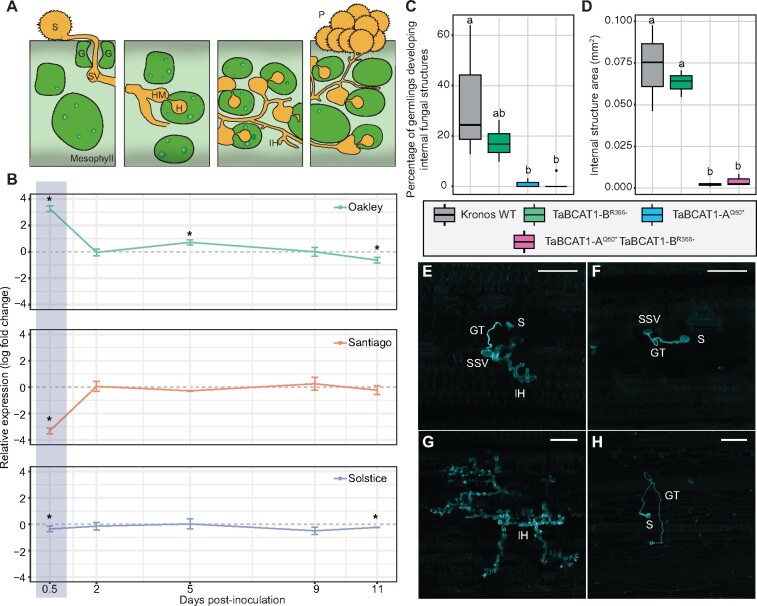
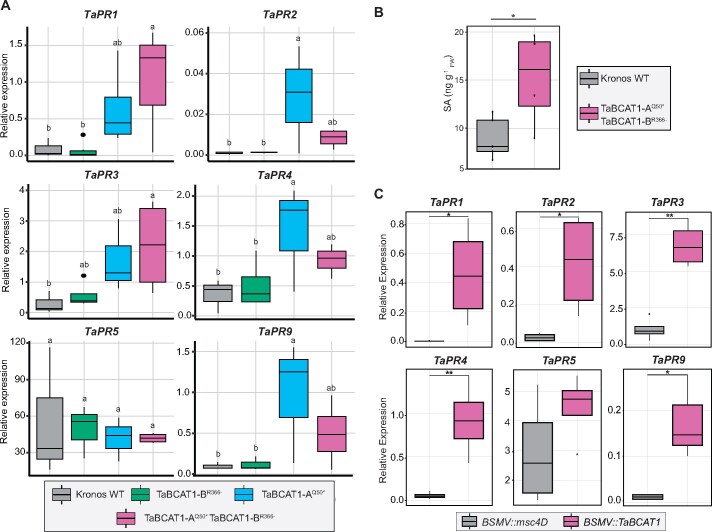
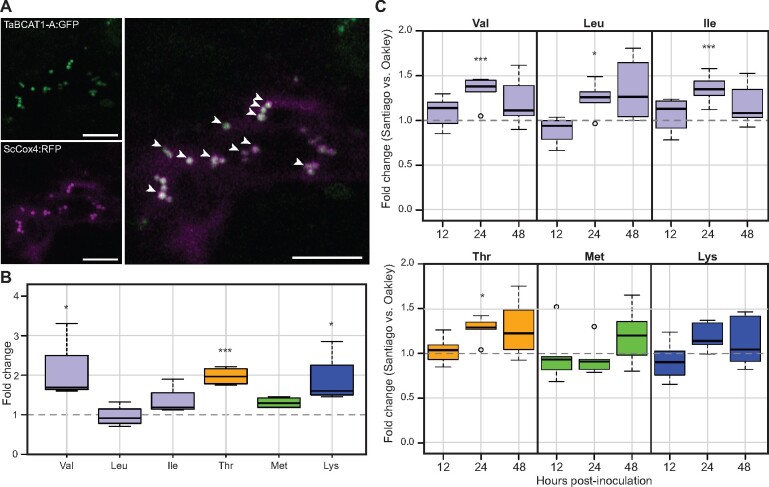
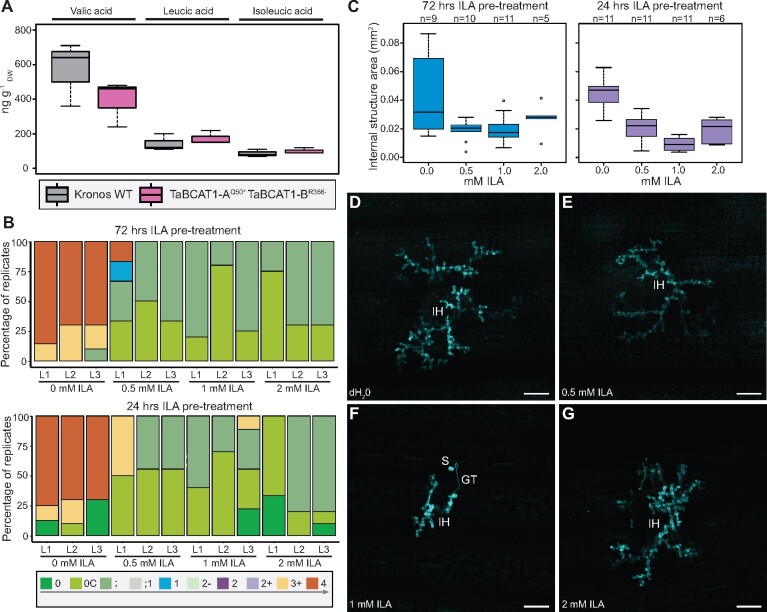
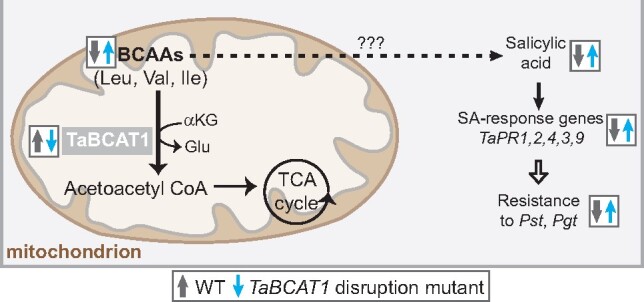
Plant pathogens suppress defense responses to evade recognition and promote successful colonization. Although identifying the genes essential for pathogen ingress has traditionally relied on screening mutant populations, the post-genomic era provides an opportunity to develop novel approaches that accelerate identification. Here, RNA-seq analysis of 68 pathogen-infected bread wheat (Triticum aestivum) varieties, including three (Oakley, Solstice and Santiago) with variable levels of susceptibility, uncovered a branched-chain amino acid aminotransferase (termed TaBCAT1) as a positive regulator of wheat rust susceptibility. We show that TaBCAT1 is required for yellow and stem rust infection and likely functions in branched-chain amino acid (BCAA) metabolism, as TaBCAT1 disruption mutants had elevated BCAA levels. TaBCAT1 mutants also exhibited increased levels of salicylic acid (SA) and enhanced expression of associated defense genes, indicating that BCAA regulation, via TaBCAT1, has a key role in SA-dependent defense activation. We also identified an association between the levels of BCAAs and resistance to yellow rust infection in wheat. These findings provide insight into SA-mediated defense responses in wheat and highlight the role of BCAA metabolism in the defense response. Furthermore, TaBCAT1 could be manipulated to potentially provide resistance to two of the most economically damaging diseases of wheat worldwide.
© The Author(s) 2021. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures


References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ (1990) Basic local alignment search tool. J Mol Biol 215: 403–410 - PubMed
-
- Bednarek P, Pislewska-Bednarek M, Svatos A, Schneider B, Doubsky J, Mansurova M, Humphry M, Consonni C, Panstruga R, Sanchez-Vallet A, et al. (2009) A glucosinolate metabolism pathway in living plant cells mediates broad-spectrum antifungal defense. Science 323: 101–106 - PubMed
-
- Bialas A, Zess EK, De la Concepcion JC, Franceschetti M, Pennington HG, Yoshida K, Upson JL, Chanclud E, Wu CH, Langner T, et al. (2018) Lessons in effector and NLR biology of plant-microbe systems. Mol Plant Microbe Interact 31: 34–45 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 715638/ERC_/European Research Council/International
- BB/M011216/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/P012574/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/P016855/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
