

Inner-nuclear-membrane-associated degradation employs Dfm1-independent retrotranslocation and alleviates misfolded transmembrane-protein toxicity
- PMID: 33566711
- PMCID: PMC8101470
- DOI: 10.1091/mbc.E20-11-0720
Inner-nuclear-membrane-associated degradation employs Dfm1-independent retrotranslocation and alleviates misfolded transmembrane-protein toxicity
Abstract
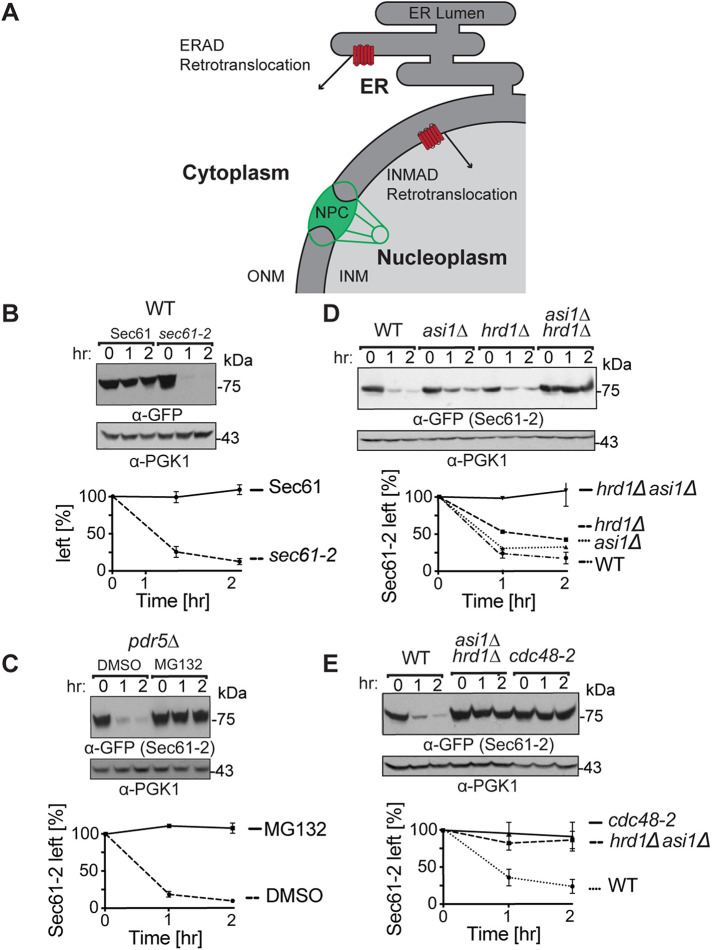
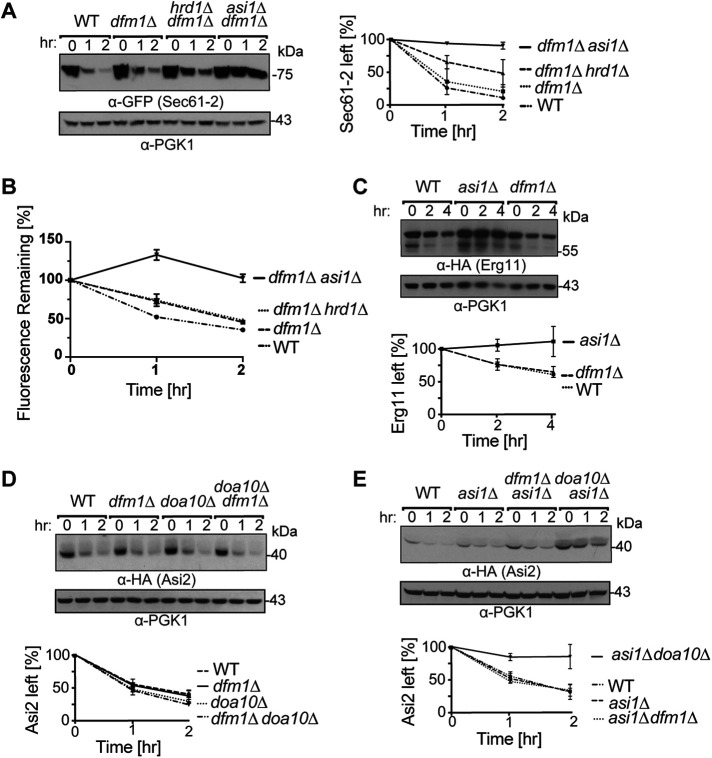
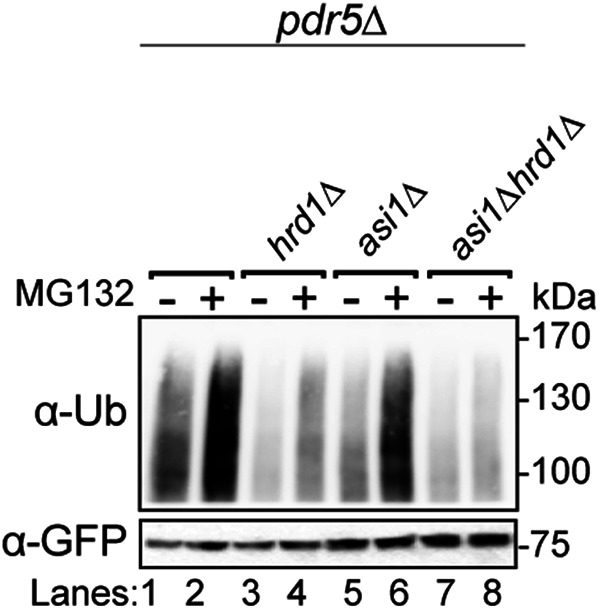
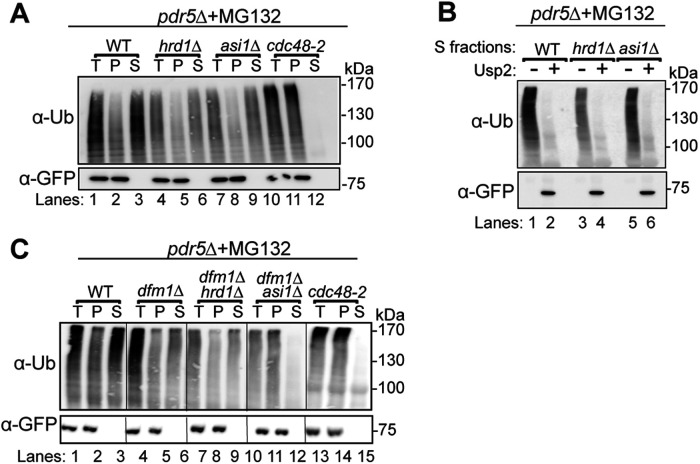
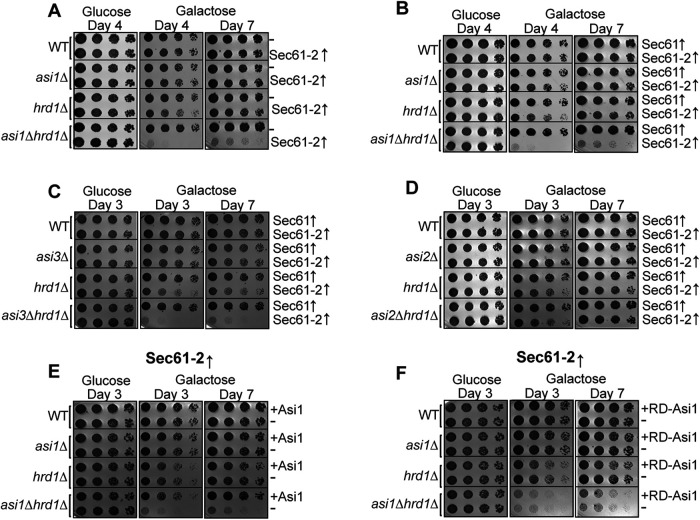
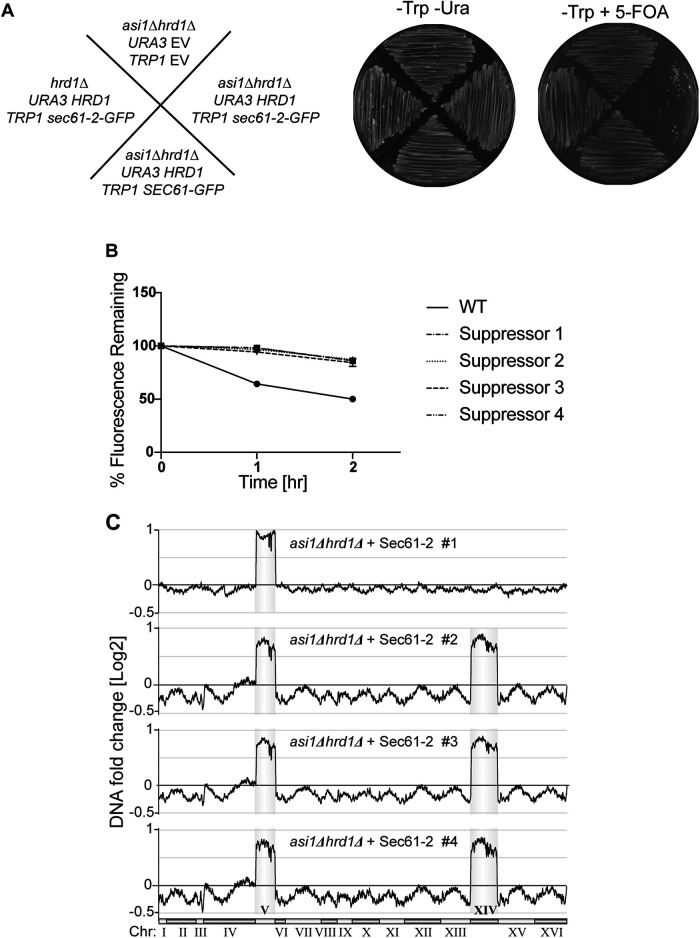
Before their delivery to and degradation by the 26S proteasome, misfolded transmembrane proteins of the endoplasmic reticulum (ER) and inner-nuclear membrane (INM) must be extracted from lipid bilayers. This extraction process, known as retrotranslocation, requires both quality-control E3 ubiquitin ligases and dislocation factors that diminish the energetic cost of dislodging the transmembrane segments of a protein. Recently, we showed that retrotranslocation of all ER transmembrane proteins requires the Dfm1 rhomboid pseudoprotease. However, we did not investigate whether Dfm1 also mediated retrotranslocation of transmembrane substrates in the INM, which is contiguous with the ER but functionally separated from it by nucleoporins. Here, we show that canonical retrotranslocation occurs during INM-associated degradation (INMAD) but proceeds independently of Dfm1. Despite this independence, ER-associated degradation (ERAD)-M and INMAD cooperate to mitigate proteotoxicity. We show a novel misfolded-transmembrane-protein toxicity that elicits genetic suppression, demonstrating the cell's ability to tolerate a toxic burden of misfolded transmembrane proteins without functional INMAD or ERAD-M. This strikingly contrasted the suppression of the dfm1Δ null, which leads to the resumption of ERAD-M through HRD-complex remodeling. Thus, we conclude that INM retrotranslocation proceeds through a novel, private channel that can be studied by virtue of its role in alleviating membrane-associated proteotoxicity.
Figures
Similar articles
-
Endoplasmic Reticulum-Associated Protein Degradation.Cold Spring Harb Perspect Biol. 2022 Dec 1;14(12):a041247. doi: 10.1101/cshperspect.a041247. Cold Spring Harb Perspect Biol. 2022. PMID: 35940909 Free PMC article. Review.
-
The Dfm1 Derlin Is Required for ERAD Retrotranslocation of Integral Membrane Proteins.Mol Cell. 2018 Jan 18;69(2):306-320.e4. doi: 10.1016/j.molcel.2017.12.012. Mol Cell. 2018. PMID: 29351849 Free PMC article.
-
Transmembrane helix hydrophobicity is an energetic barrier during the retrotranslocation of integral membrane ERAD substrates.Mol Biol Cell. 2017 Jul 15;28(15):2076-2090. doi: 10.1091/mbc.E17-03-0184. Epub 2017 May 24. Mol Biol Cell. 2017. PMID: 28539401 Free PMC article.
-
Cdc48 and Ubx1 participate in a pathway associated with the inner nuclear membrane that governs Asi1 degradation.J Cell Sci. 2016 Oct 15;129(20):3770-3780. doi: 10.1242/jcs.189332. Epub 2016 Aug 26. J Cell Sci. 2016. PMID: 27566164
-
Towards understanding inner nuclear membrane protein degradation in plants.J Exp Bot. 2022 Apr 18;73(8):2266-2274. doi: 10.1093/jxb/erac037. J Exp Bot. 2022. PMID: 35139191 Review.
Cited by
-
Endoplasmic Reticulum-Associated Protein Degradation.Cold Spring Harb Perspect Biol. 2022 Dec 1;14(12):a041247. doi: 10.1101/cshperspect.a041247. Cold Spring Harb Perspect Biol. 2022. PMID: 35940909 Free PMC article. Review.
-
Elements of the ERAD ubiquitin ligase Doa10 regulating sequential poly-ubiquitylation of its targets.iScience. 2022 Oct 13;25(11):105351. doi: 10.1016/j.isci.2022.105351. eCollection 2022 Nov 18. iScience. 2022. PMID: 36325070 Free PMC article.
-
SLC26A9 is selected for endoplasmic reticulum associated degradation (ERAD) via Hsp70-dependent targeting of the soluble STAS domain.Biochem J. 2021 Dec 22;478(24):4203-4220. doi: 10.1042/BCJ20210644. Biochem J. 2021. PMID: 34821356 Free PMC article.
-
Inner Nuclear Membrane Asi Ubiquitin Ligase Catalytic Subunits Asi1p and Asi3p, but not Asi2p, confer resistance to aminoglycoside hygromycin B in Saccharomyces cerevisiae.MicroPubl Biol. 2021 Jun 1;2021:10.17912/micropub.biology.000403. doi: 10.17912/micropub.biology.000403. MicroPubl Biol. 2021. PMID: 34095778 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
