

VviNAC33 promotes organ de-greening and represses vegetative growth during the vegetative-to-mature phase transition in grapevine
- PMID: 33567124
- PMCID: PMC8251598
- DOI: 10.1111/nph.17263
VviNAC33 promotes organ de-greening and represses vegetative growth during the vegetative-to-mature phase transition in grapevine
Abstract
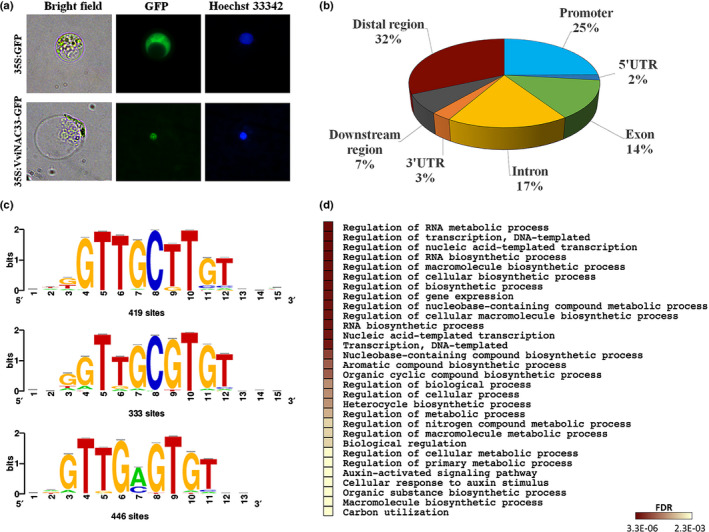
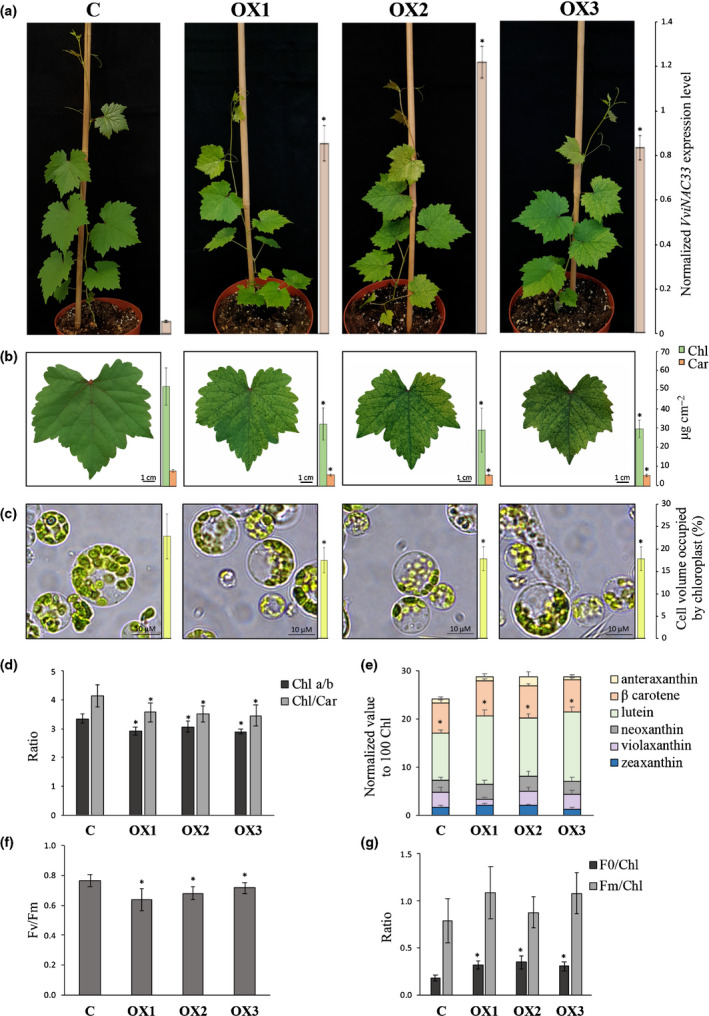
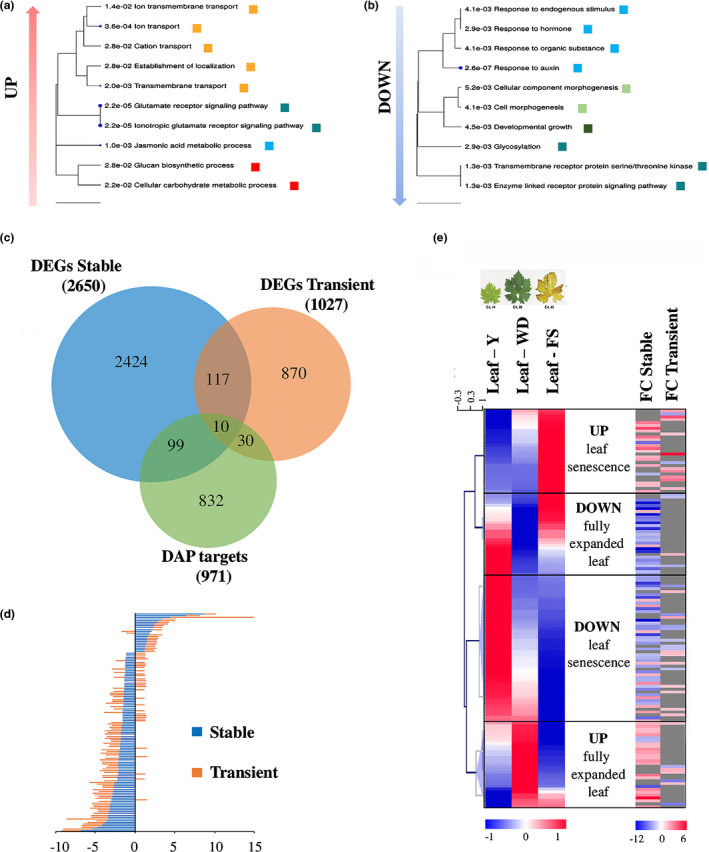
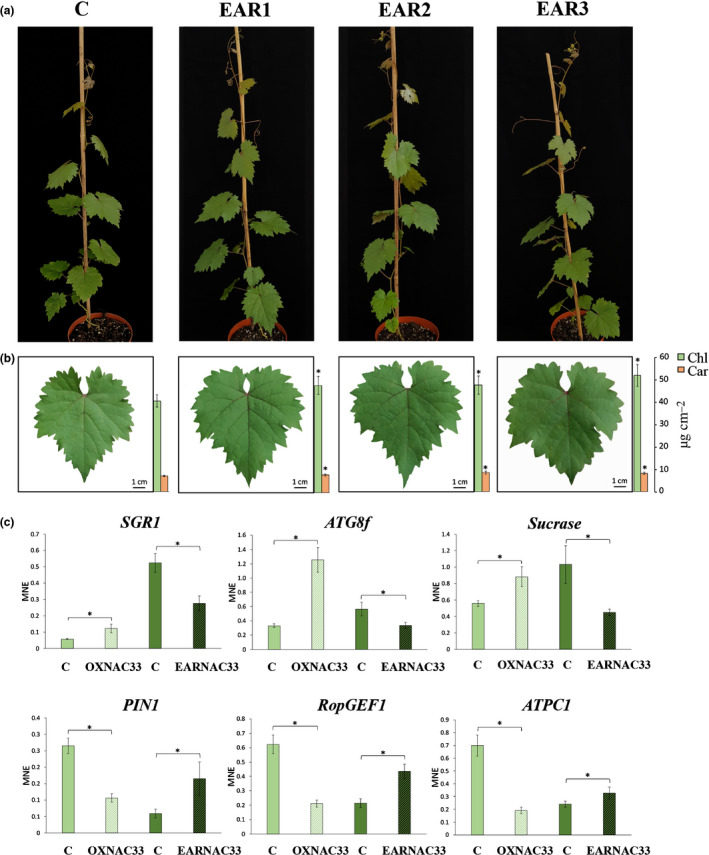
Plants undergo several developmental transitions during their life cycle. In grapevine, a perennial woody fruit crop, the transition from vegetative/green-to-mature/woody growth involves transcriptomic reprogramming orchestrated by a small group of genes encoding regulators, but the underlying molecular mechanisms are not fully understood. We investigated the function of the transcriptional regulator VviNAC33 by generating and characterizing transgenic overexpressing grapevine lines and a chimeric repressor, and by exploring its putative targets through a DNA affinity purification sequencing (DAP-seq) approach combined with transcriptomic data. We demonstrated that VviNAC33 induces leaf de-greening, inhibits organ growth and directly activates the expression of STAY-GREEN PROTEIN 1 (SGR1), which is involved in Chl and photosystem degradation, and AUTOPHAGY 8f (ATG8f), which is involved in the maturation of autophagosomes. Furthermore, we show that VviNAC33 directly inhibits AUXIN EFFLUX FACILITATOR PIN1, RopGEF1 and ATP SYNTHASE GAMMA CHAIN 1T (ATPC1), which are involved in photosystem II integrity and activity. Our results show that VviNAC33 plays a major role in terminating photosynthetic activity and organ growth as part of a regulatory network governing the vegetative-to-mature phase transition.
Keywords: DAP-seq; NAC33; de-greening; grapevine; phase transition; transcriptomics; vegetative growth.
© 2021 The Authors New Phytologist © 2021 New Phytologist Foundation.
Figures




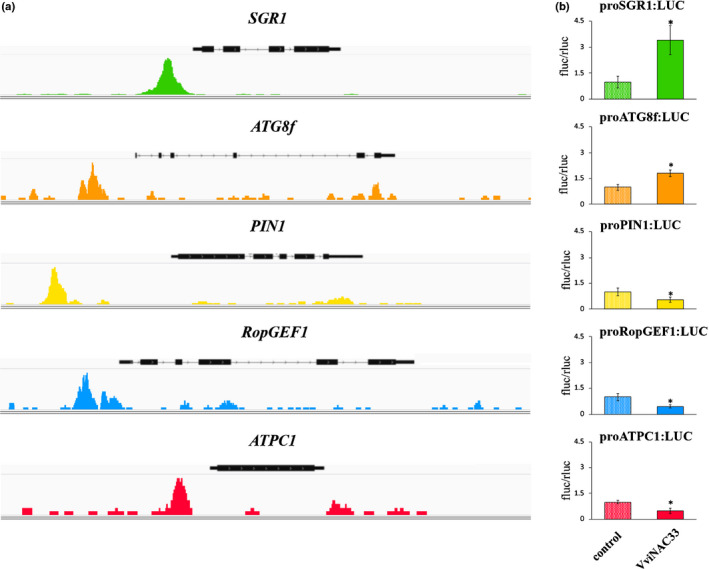
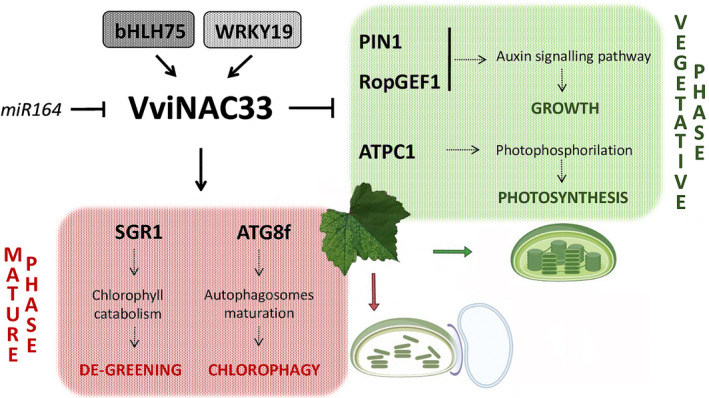
Comment in
-
Time to retire? A life-changing decision made by NAC transcription factors.New Phytol. 2021 Jul;231(2):505-507. doi: 10.1111/nph.17451. New Phytol. 2021. PMID: 34132413 No abstract available.
References
-
- Amato A, Cavallini E, Walker AR, Pezzotti M, Bliek M, Quattrocchio F, Koes R, Ruperti B, Bertini E, Zenoni S et al. 2019. The MYB5‐driven MBW complex recruits a WRKY factor to enhance the expression of targets involved in vacuolar hyper‐acidification and trafficking in grapevine. The Plant Journal 99: 1220–1241. - PubMed
-
- Baker NR. 2008. Chlorophyll fluorescence: a probe of photosynthesis in vivo. Annual Review of Plant Biology 59: 89–113. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
